Siderocalin
| Neutrophil gelatinase-associated lipocalin/epididymal-specific lipocalin-12 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Aliases | LCN2/LCN12IPR003087Siderocalin | ||||||
| External IDs | GeneCards: [1] | ||||||
| Orthologs | |||||||
| Species | Human | Mouse | |||||
| Entrez |
|
| |||||
| Ensembl |
|
| |||||
| UniProt |
| ||||||
| RefSeq (mRNA) |
|
| |||||
| RefSeq (protein) |
|
| |||||
| Location (UCSC) | n/a | n/a | |||||
| PubMed search | n/a | n/a | |||||
| Wikidata | |||||||
| |||||||
Siderocalin (Scn), lipocalin-2, NGAL, 24p3 is a mammalian lipocalin-type protein that can prevent iron acquisition by pathogenic bacteria by binding siderophores, which are iron-binding chelators made by microorganisms.[1][2] Iron serves as a key nutrient in host-pathogen interactions, and pathogens can acquire iron from the host organism via synthesis and release siderophores such as enterobactin.[3] Siderocalin is a part of the mammalian defence mechanism and acts as an antibacterial agent.[1][4][5][6][7] Crystallographic studies of Scn demonstrated that it includes a calyx, a ligand-binding domain that is lined with polar cationic groups.[8] Central to the siderophore/siderocalin recognition mechanism are hybrid electrostatic/cation-pi interactions.[5][9] To evade the host defences, pathogens evolved to produce structurally varied siderophores that would not be recognized by siderocalin, allowing the bacteria to acquire iron.[1]
Iron requirements of host organisms[]
Organisms require iron for a variety of chemical reactions.[10] Although iron can be found throughout the biosphere, free ferric iron forms insoluble hydroxides at physiological pH, limiting its accessibility in aerobic conditions to living organisms.[10][11] In order to preserve homeostasis, organisms have evolved specific protein networks, with proteins and receptors translated in accordance with intracellular iron levels.[10][12] Export and import are supplemented by a cycling process between the ferrous Fe(II) available in the reducing environment of the cell, and ferric Fe(III) found primarily under aerobic conditions.[13] [14] The iron acquisition mechanisms of pathogenic bacteria demonstrate the role of iron as a key component at the interface between pathogens and hosts.[13][14]
Lipocalin family of iron binding proteins[]
The lipocalin family of binding proteins are produced by the immune system and sequester ferric siderophore complexes from the siderophore receptors of bacteria.[15] [16] The lipocalin family of binding proteins typically have a conserved eight-stranded β-barrel fold with a calyx binding site,[16][17] which are lined with positively charged amino acid residues, allowing for binding interactions with siderophores.[citation needed]
Clinical significance[]
Mycobacterial infections[]
The lipocalin siderocalin is found in neutrophil granules, uterine secretions, and at particularly high levels in serum during bacterial infection.[4] Upon infection, pathogens use siderophores to capture iron from the host organism.[18] This strategy is, however, complicated by the human protein siderocalin, which can sequester siderophores, and prevent their use by pathogenic bacteria as iron delivery agents.[19] This effect has been demonstrated by studies with siderocalin-knock-out mice, which are more sensitive to infections under iron-limiting conditions.[4][5]
Mycobacterial virulence[]
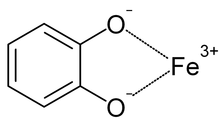
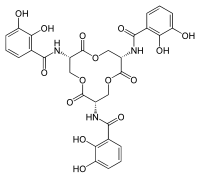
Siderophores are iron chelators, allowing organisms to acquire iron from their environment. In the case of pathogens, iron can be acquired from the host organism.[20] Siderophores and ferric iron can associate to form stable complexes.[10][21][22] Siderophores bind iron using a variety of ligands, most commonly as α-hydroxycarboxylates (e.g. citrate), catecholates, and hydroxamates.[5][10][23] [24] As a defence mechanism, siderocalin can substitute ferric bis-catechol complexes (formed under physiological conditions) with a third catechol, in order to achieve a hexacoordinate ferric complex, resulting in higher affinity binding.[5][18][25]
As a mediator of mammalian iron transport[]
Mammalian siderophores, specifically catechols, can be found in the human gut and in siderophores, such as enterobactin, and serve as iron-binding moieties.[5][26] Catechol resembling molecules can act as iron ligands in the cell and in systematic circulation, allowing siderocalin to bind to the iron-catechol complex.[27] Catechols can be bound by siderocalin, in the form of free ligands, or in the iron complex.[28] 24p3 is a vertebrate lipocalin-2 receptor which allows for import of the ferric siderophore complex into mammalian cells.[27] During kidney embryogenesis, siderocalin mediated iron transport occurs, as iron concentration has to be highly controlled in order to restrict inflammation.[4][11] Following secretion by neutrophils, siderocalin can bind to pathogenic siderophores, such as bacillibactin, and prevent siderophore trafficking.[29] Siderocalin has been linked with various cellular processes apart from iron transport, including apoptosis, cellular differentiation, tumorigenesis, and metastasis.[10][30]
Structure[]
The avian orthologs of siderocalin (Q83 and Ex-FABP) and NGAL (neutrophil gelatinase-associated lipocalin-2) contain calyces with positively charged lysine and arginine side chains.[8][30][31][32][33] These side chains interact via cation-pi and coulombic interactions with the negatively charged siderophores that contain aromatic catecholate groups.[10][30] Crystallographic studies of siderocalin have shown that the ligand binding domain of Scn, known as the calyx, is shallow and broad, and is lined with polar cationic groups from the three positively charged residues of Arg81, Lys125, and Lys134.[5][8][34] Scn can also bind non-ferric complexes and has been identified as a potential transporter for heavy actinide ions. Scn crystal structures containing heavy metals (thorium, plutonium, americium, curium, and californium) have been obtained.[35][36] Scn has been found as a monomer, homo-dimer, or trimer in human plasma.[5] The siderocalin fold is exceptionally stable.[4][5] The calyx is structurally stable and rigid, and conformational change does not typically occur upon a change in pH, ionic strength, or ligand binding.[5]
Binding pocket[]
The structural stability of the calyx has been attributed to the three binding pockets within the calyx that sterically limit which ligands are compatible with siderocalin.[5][8] The Scn calyx can accommodate three aromatic rings of the catecholate moieties, in the three available binding pockets.[5][28] Solid-state and solution structural results demonstrated that bacteria-derived enterobactin is bound to the binding pocket of Scn, allowing for Scn to be involved in the acute immune response to bacterial infection.[5][21] One method by which pathogens can circumvent immunity mechanisms is by modifying the siderophore chemical structure to prevent interaction with Scn.[24] One example is the addition of glucose molecules to the enterobactin backbone of salmochelin (C-glucosylated enterobactin) in order to increase the hydrophilicity and bulkiness of a siderophore and inhibit binding to Scn.[24][37]
Binding interactions[]
Siderophores are typically bound to siderocalin with subnanomolar affinities, and interact with siderocalin specifically.[10][25] The Kd value of the siderocalin/siderophore interaction, measured by fluorescence quenching (Kd= 0.4 nM), indicates that siderocalin can capture siderophores with high affinity.[31][38] This Kd value is similar to that of the FepA bacterial receptor (Kd= 0.3 nM).[5] Siderophore/siderocalin binding is directed by electrostatic interactions.[5][38] Specifically, the mechanism involves hybrid electrostatic and cation-pi interactions in the positively charged protein calyx.[25] The siderophore is positioned in the centre of the siderocalin calyx, and is associated with multiple direct polar interactions.[25] Structural analysis of the siderocalin/siderophore interaction has shown that the siderophore is accompanied by a poor and diffuse quality of electron density, with the majority of the ligand exposed to the solvent when the siderophore is fit in the calyx.[5][6] Siderocalin typically does not bind hydroxamate-based siderophores because these substrates do not have the necessary aromatic electronic structure for cation-pi interactions.[5][25] In order to acquire iron in the presence of siderocalin, pathogenic bacteria utilize several siderophores that do not bind to siderocalin, or structurally modify siderophores to inhibit siderocalin binding.[5][39] Siderocalin can bind soluble siderophores of mycobacteria, including carboxymycobactins.[5][6] In vivo studies have shown that the binding interactions between carboxymycobactin and siderocalin serve to protect the host organism from mycobacterial infections, with siderocalin inhibiting mycobacterial iron acquisition.[5][28][40] Siderocalin can sequester ferric carboxymycobactins by employing a polyspecific recognition mechanism.[5] The siderophore/siderocalin recognition mechanism primarily involves hybrid electrostatic/cation-pi interactions.[5][9][11] The fatty acid tails of carboxymycobactin reside in a ‘tail-in’ or ‘tail-out’ conformation within pocket 2.[5] The ‘tail-in’ conformation of the fatty acid chain lengths introduces a significant interaction between the calyx and the ligand, increasing the affinity of the siderocalin calyx and carboxymycobactin.[5] The fatty acid tails of short lengths have a correspondingly less favorable binding to siderocalin, and cannot maintain the necessary interaction with the binding pocket.[5] Since lipocalin-2 cannot bind the long fatty acid chain carboxymycobactins of mycobacteria, it is apparent that a number of pathogens have evolved to avoid the activity of lipocalin-2.[41]
Recognition mechanism[]
Electrostatic interaction play a key role in the recognition mechanism of siderophores by siderocalin.[1] The binding of the siderophore and the siderocalin binding pocket is primarily directed by cation-pi interactions, with the positively charged binding pocket of siderocalin attracting the negatively charged complex.[1] A structural factor involved in the siderocalin mediated recognition mechanism of phenolate/catecholate-type siderophores includes a backbone linker which allows for siderocalin to interact with different phenolate/catecholate siderophores.[4][42] While siderocalin recognition is minimally affected by the substitution of different metals, methylating the three catecholate rings of enterobactin can impede the recognition of siderocalin.[5][34][38][43] A strategy used by pathogens to overcome immune response is the production of siderophores that will not be recognized by siderocalin.[19][44] For example, siderocalin cannot recognize the siderophores of the C-glucosylated analog of enterobactin, as the donor groups are glycosylated, introducing steric interactions at the position 5-carbons of the catechol groups.[1][24]
History[]
The requirement for iron by humans and pathogens has been known for many years.[10] The link between iron and mycobactins, iron-chelating growth factors from mycobacteria, was first made in the 1960s.[5] At the time, interest was growing in resolving an application of mycobactins as target molecules for a rational anti-tuberculosis agent.[5][45] Experiments in the 1960s and 1970s demonstrated that iron deficiency in mycobacteria was the cause of 'anaemic’ cells.[46] The majority of the genes and systems necessary for high affinity iron acquisition have been identified in pathogenic and saprophytic mycobacteria.[5] These genes encode proteins for iron storage, uptake of ferric-siderophores, and heme.[5][47] Humans have evolved a defense for siderophore-mediated iron acquisition by developing siderocalin. To combat this, various pathogens have evolved siderophores that can evade siderocalin recognition.[5] Siderocalin has been shown to bind to siderophores and inhibit iron acquisition, and prevent the growth of Mycobacterium tuberculosis in extracellular cultures; however, the effect of siderocalin on this pathogen within macrophages remains unclear.[24][31]
See also[]
- LCN2
- LCN1
- Animal pathogens
- Mycobacteria
References[]
- ^ Jump up to: a b c d e f van Eldik R, Hubbard CD (2009). Advances in Inorganic Chemistry Volume 61 (1st ed.). London, U.K.: Elsevier. pp. 237–239. ISBN 9780123750334. Retrieved 16 February 2015.
- ^ Correnti C, Richardson V, Sia AK, Bandaranayake AD, Ruiz M, Suryo Rahmanto Y, Kovačević Ž, Clifton MC, Holmes MA, Kaiser BK, Barasch J, Raymond KN, Richardson DR, Strong RK (2012). "Siderocalin/Lcn2/NGAL/24p3 does not drive apoptosis through gentisic acid mediated iron withdrawal in hematopoietic cell lines". PLOS ONE. 7 (8): e43696. Bibcode:2012PLoSO...743696C. doi:10.1371/journal.pone.0043696. PMC 3424236. PMID 22928018.
- ^ Cherayil BJ (May 2011). "The role of iron in the immune response to bacterial infection". Immunologic Research. 50 (1): 1–9. doi:10.1007/s12026-010-8199-1. PMC 3085559. PMID 21161695.
- ^ Jump up to: a b c d e f Paragas N, Qiu A, Hollmen M, Nickolas TL, Devarajan P, Barasch J (Sep 2012). "NGAL-Siderocalin in kidney disease". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1823 (9): 1451–8. doi:10.1016/j.bbamcr.2012.06.014. PMC 3664277. PMID 22728330.
- ^ Jump up to: a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae Byers BR (2013). Iron Acquisition by the Genus Mycobacterium. SpringerBriefs in Molecular Science. Springer. pp. 1–88. doi:10.1007/978-3-319-00303-0. ISBN 978-3-319-00303-0. S2CID 12666634.
- ^ Jump up to: a b c Holmes MA, Paulsene W, Jide X, Ratledge C, Strong RK (Jan 2005). "Siderocalin (Lcn 2) also binds carboxymycobactins, potentially defending against mycobacterial infections through iron sequestration". Structure. 13 (1): 29–41. doi:10.1016/j.str.2004.10.009. PMID 15642259.
- ^ Sige l A, Sigel H, Sige l RK (2013). Interrelations between Essential Metal Ions and Human Diseases. Heidelberg, Germany: Springer. pp. 282–283. ISBN 9789400774995. Retrieved 14 February 2015.
- ^ Jump up to: a b c d Sia AK, Allred BE, Raymond KN (Apr 2013). "Siderocalins: Siderophore binding proteins evolved for primary pathogen host defense". Current Opinion in Chemical Biology. 17 (2): 150–7. doi:10.1016/j.cbpa.2012.11.014. PMC 3634885. PMID 23265976.
- ^ Jump up to: a b Abergel RJ, Wilson MK, Arceneaux JE, Hoette TM, Strong RK, Byers BR, Raymond KN (Dec 2006). "Anthrax pathogen evades the mammalian immune system through stealth siderophore production". Proceedings of the National Academy of Sciences of the United States of America. 103 (49): 18499–503. Bibcode:2006PNAS..10318499A. doi:10.1073/pnas.0607055103. PMC 1693691. PMID 17132740.
- ^ Jump up to: a b c d e f g h i Correnti C, Strong RK (Apr 2012). "Mammalian siderophores, siderophore-binding lipocalins, and the labile iron pool". The Journal of Biological Chemistry. 287 (17): 13524–31. doi:10.1074/jbc.R111.311829. PMC 3340207. PMID 22389496.
- ^ Jump up to: a b c Chakraborty R, Braun V, Hantke K, Cornelis P (2013). Iron Uptake in Bacteria with Emphasis on E. coli and Pseudomonas. SpringerBriefs in Biometals. pp. 31–66. ISBN 978-94-007-6087-5.
- ^ Ganz T (Oct 2013). "Systemic iron homeostasis". Physiological Reviews. 93 (4): 1721–41. doi:10.1152/physrev.00008.2013. PMID 24137020.
- ^ Jump up to: a b Doherty CP (May 2007). "Host-pathogen interactions: the role of iron". The Journal of Nutrition. 137 (5): 1341–4. doi:10.1093/jn/137.5.1341. PMID 17449603.
- ^ Jump up to: a b Skaar EP (2010). "The battle for iron between bacterial pathogens and their vertebrate hosts". PLOS Pathogens. 6 (8): e1000949. doi:10.1371/journal.ppat.1000949. PMC 2920840. PMID 20711357.
- ^ Sandy M, Butler A (Oct 2009). "Microbial iron acquisition: marine and terrestrial siderophores". Chemical Reviews. 109 (10): 4580–95. doi:10.1021/cr9002787. PMC 2761978. PMID 19772347.
- ^ Jump up to: a b Flower DR (Aug 1996). "The lipocalin protein family: structure and function". The Biochemical Journal. 318 (1): 1–14. doi:10.1042/bj3180001. PMC 1217580. PMID 8761444.
- ^ Fuentes-Prior P, Noeske-Jungblut C, Donner P, Schleuning WD, Huber R, Bode W (Oct 1997). "Structure of the thrombin complex with triabin, a lipocalin-like exosite-binding inhibitor derived from a triatomine bug". Proceedings of the National Academy of Sciences of the United States of America. 94 (22): 11845–50. Bibcode:1997PNAS...9411845F. doi:10.1073/pnas.94.22.11845. PMC 23629. PMID 9342325.
- ^ Jump up to: a b Miethke M, Marahiel MA (Sep 2007). "Siderophore-based iron acquisition and pathogen control". Microbiology and Molecular Biology Reviews. 71 (3): 413–51. doi:10.1128/MMBR.00012-07. PMC 2168645. PMID 17804665.
- ^ Jump up to: a b Allred BE, Correnti C, Clifton MC, Strong RK, Raymond KN (Sep 2013). "Siderocalin outwits the coordination chemistry of vibriobactin, a siderophore of Vibrio cholerae". ACS Chemical Biology. 8 (9): 1882–7. doi:10.1021/cb4002552. PMC 3783644. PMID 23755875.
- ^ Miethke M (Jan 2013). "Molecular strategies of microbial iron assimilation: from high-affinity complexes to cofactor assembly systems". Metallomics. 5 (1): 15–28. doi:10.1039/C2MT20193C. PMID 23192658.
- ^ Jump up to: a b Abergel RJ, Clifton MC, Pizarro JC, Warner JA, Shuh DK, Strong RK, Raymond KN (Aug 2008). "The siderocalin/enterobactin interaction: a link between mammalian immunity and bacterial iron transport". Journal of the American Chemical Society. 130 (34): 11524–34. doi:10.1021/ja803524w. PMC 3188318. PMID 18680288.
- ^ Fukushima T, Allred BE, Sia AK, Nichiporuk R, Andersen UN, Raymond KN (Aug 2013). "Gram-positive siderophore-shuttle with iron-exchange from Fe-siderophore to apo-siderophore by Bacillus cereus YxeB". Proceedings of the National Academy of Sciences of the United States of America. 110 (34): 13821–6. Bibcode:2013PNAS..11013821F. doi:10.1073/pnas.1304235110. PMC 3752266. PMID 23924612.
- ^ Dhungana S, Harrington JM, Gebhardt P, Möllmann U, Crumbliss AL (Oct 2007). "Iron chelation equilibria, redox, and siderophore activity of a saccharide platform ferrichrome analogue" (PDF). Inorganic Chemistry. 46 (20): 8362–71. doi:10.1021/ic070158l. PMID 17824601.
- ^ Jump up to: a b c d e Yehuda S, Mostofsky DI (2010). Iron Deficiency and Overload: From Basic Biology to Clinical Medicine. New York, N.Y.: Humana Press. pp. 66–69. ISBN 9781934115220. Retrieved 14 February 2015.
- ^ Jump up to: a b c d e Hoette TM, Abergel RJ, Xu J, Strong RK, Raymond KN (Dec 2008). "The role of electrostatics in siderophore recognition by the immunoprotein Siderocalin". Journal of the American Chemical Society. 130 (51): 17584–92. doi:10.1021/ja8074665. PMC 2778733. PMID 19053425.
- ^ Rogers HJ (Mar 1973). "Iron-Binding Catechols and Virulence in Escherichia coli". Infection and Immunity. 7 (3): 445–56. doi:10.1128/IAI.7.3.445-456.1973. PMC 422698. PMID 16558077.
- ^ Jump up to: a b Anderson GJ, McLaren GD (2012). Iron Physiology and Pathophysiology in Humans. New York, N.Y.: Springer. pp. 237–239, 658. ISBN 9781603274845. Retrieved 14 February 2015.
- ^ Jump up to: a b c Bao G, Clifton M, Hoette TM, Mori K, Deng SX, Qiu A, Viltard M, Williams D, Paragas N, Leete T, Kulkarni R, Li X, Lee B, Kalandadze A, Ratner AJ, Pizarro JC, Schmidt-Ott KM, Landry DW, Raymond KN, Strong RK, Barasch J (Aug 2010). "Iron traffics in circulation bound to a siderocalin (Ngal)-catechol complex". Nature Chemical Biology. 6 (8): 602–9. doi:10.1038/nchembio.402. PMC 2907470. PMID 20581821.
- ^ Bergman NH (2011). Bacillus anthracis and Anthrax. Hoboken, N.J.: Wiley. pp. Chapter 7. ISBN 9781118148082. Retrieved 14 February 2015.
- ^ Jump up to: a b c Correnti C, Clifton MC, Abergel RJ, Allred B, Hoette TM, Ruiz M, Cancedda R, Raymond KN, Descalzi F, Strong RK (Dec 2011). "Galline Ex-FABP is an antibacterial siderocalin and a lysophosphatidic acid sensor functioning through dual ligand specificities". Structure. 19 (12): 1796–806. doi:10.1016/j.str.2011.09.019. PMC 3240821. PMID 22153502.
- ^ Jump up to: a b c Ashton Acton Q (2012). Advances in Serine Research and Application (2012: ScholarlyBrief ed.). Atlanta, Georgia: ScholarlyEditions. pp. 42–43. ISBN 9781481614276. Retrieved 14 February 2015.
- ^ Thongboonkerd V (2007). Proteomics of Human Body Fluids: Principles, Methods, and Applications. Totowa, N.J: Humana Press. pp. 338–339. ISBN 9781597454322. Retrieved 16 February 2015.
- ^ Clifton MC, Corrent C, Strong RK (Aug 2009). "Siderocalins: siderophore-binding proteins of the innate immune system" (PDF). Biometals. 22 (4): 557–64. doi:10.1007/s10534-009-9207-6. PMID 19184458. S2CID 8776376.
- ^ Jump up to: a b Hoette TM, Clifton MC, Zawadzka AM, Holmes MA, Strong RK, Raymond KN (Dec 2011). "Immune interference in Mycobacterium tuberculosis intracellular iron acquisition through siderocalin recognition of carboxymycobactins". ACS Chemical Biology. 6 (12): 1327–31. doi:10.1021/cb200331g. PMC 3241878. PMID 21978368.
- ^ Deblonde, Gauthier J.-P.; Sturzbecher-Hoehne, Manuel; Rupert, Peter B.; Dahlia, An D.; Illy Marie-Claire; Ralston, Corie Y.; Brabec, Jiri; de Jong, Wide A.; Strong, Roland (September 2017). "Chelation and stabilization of berkelium in oxidation state +IV" (PDF). Nature Chemistry. 9 (9): 843–849. Bibcode:2017NatCh...9..843D. doi:10.1038/nchem.2759. ISSN 1755-4349. OSTI 1436161. PMID 28837177.
- ^ Captain, Ilya; Deblonde, Gauthier J.-P.; Rupert, Peter B.; An, Dahlia D.; Illy, Marie-Claire; Rostan, Emeline; Ralston, Corie Y.; Strong, Roland K.; Abergel, Rebecca J. (2016-11-21). "Engineered Recognition of Tetravalent Zirconium and Thorium by Chelator–Protein Systems: Toward Flexible Radiotherapy and Imaging Platforms". Inorganic Chemistry. 55 (22): 11930–11936. doi:10.1021/acs.inorgchem.6b02041. ISSN 0020-1669. OSTI 1458481. PMID 27802058.
- ^ Alvarez MV (2007). Isolation, Structure and Detection of Salmochelins: Novel Siderophores in Enterobacteria. Göttingen, Germany: Cuvillier Verlag. pp. 29–34. ISBN 9783867271097. Retrieved 14 February 2015.
- ^ Jump up to: a b c Abergel RJ, Moore EG, Strong RK, Raymond KN (Aug 2006). "Microbial evasion of the immune system: structural modifications of enterobactin impair siderocalin recognition". Journal of the American Chemical Society. 128 (34): 10998–9. doi:10.1021/ja062476+. PMC 3188317. PMID 16925397.
- ^ Allela L, Boury O, Pouillot R, Délicat A, Yaba P, Kumulungui B, Rouquet P, Gonzalez JP, Leroy EM (Mar 2005). "Ebola virus antibody prevalence in dogs and human risk". Emerging Infectious Diseases. 11 (3): 385–90. doi:10.3201/eid1103.040981. PMC 3298261. PMID 15757552.
- ^ Åkerström B (2006). Lipocalins. Austin, Texas: Landes Bioscience. p. 92. ISBN 9781587062971.
- ^ Kidd SP (2011). Stress Response in Pathogenic Bacteria, Volume 19 of Advances in Molecular and Cellular Microbiology. Wallingford, U.K.: CABI. pp. 287–290. ISBN 9781845937775. Retrieved 14 February 2015.
- ^ Strong, R. K.; Akerstrom, B.; Borregaard, N.; Flower, D. R.; Salier, J.-P. (Eds.). "Siderocalins" (PDF). Fred Hutchinson Cancer Research Center.CS1 maint: extra text: authors list (link)
- ^ Abergel RJ, Warner JA, Shuh DK, Raymond KN (Jul 2006). "Enterobactin protonation and iron release: structural characterization of the salicylate coordination shift in ferric enterobactin". Journal of the American Chemical Society. 128 (27): 8920–31. doi:10.1021/ja062046j. PMC 3188320. PMID 16819888.
- ^ Stintzi A, Barnes C, Xu J, Raymond KN (Sep 2000). "Microbial iron transport via a siderophore shuttle: a membrane ion transport paradigm". Proceedings of the National Academy of Sciences of the United States of America. 97 (20): 10691–6. Bibcode:2000PNAS...9710691S. doi:10.1073/pnas.200318797. PMC 27084. PMID 10995480.
- ^ Barclay R, Ratledge C (Mar 1983). "Iron-binding compounds of Mycobacterium avium, M. intracellulare, M. scrofulaceum, and mycobactin-dependent M. paratuberculosis and M. avium". Journal of Bacteriology. 153 (3): 1138–46. doi:10.1128/JB.153.3.1138-1146.1983. PMC 221756. PMID 6826517.
- ^ Jamison DT, Breman JG, Measham AR, Alleyne G, Claeson M, Evans DB, Jha P, Mills A, Musgrove P (2006). Disease Control Priorities in Developing Countries (2nd ed.). Washington, D.C.: World Bank. pp. Chapter 16. ISBN 978-0-8213-6179-5. Retrieved 16 February 2015.
- ^ Caza M, Kronstad JW (2013). "Shared and distinct mechanisms of iron acquisition by bacterial and fungal pathogens of humans". Frontiers in Cellular and Infection Microbiology. 3 (80): 80. doi:10.3389/fcimb.2013.00080. PMC 3832793. PMID 24312900.
- Genes on human chromosome
- Proteins