Common buzzard
| Common buzzard | |
|---|---|

| |
| Steppe buzzard B. b. vulpinus | |
Conservation status
| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Aves |
| Order: | Accipitriformes |
| Family: | Accipitridae |
| Genus: | Buteo |
| Species: | B. buteo
|
| Binomial name | |
| Buteo buteo (Linnaeus, 1758)
| |
| Subspecies | |
|
7–10, see text | |
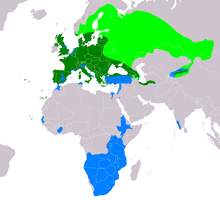
| |
| Range of B. buteo Breeding range of migrant populations Resident range Wintering range of migrant populations
| |
| Synonyms | |
|
Falco buteo Linnaeus, 1758 | |

The common buzzard (Buteo buteo) is a medium-to-large bird of prey which has a large range. A member of the genus Buteo, it is a member of the family Accipitridae. The species lives in most of Europe and extends its breeding range across much of the Palearctic as far as the northwestern China (Tian Shan), far western Siberia and northwestern Mongolia.[1][2] Over much of its range, it is a year-round resident. However, buzzards from the colder parts of the Northern Hemisphere as well as those that breed in the eastern part of their range typically migrate south for the northern winter, many journeying as far as South Africa.[3] The common buzzard is an opportunistic predator that can take a wide variety of prey, but it feeds mostly on small mammals, especially rodents such as voles. It typically hunts from a perch.[4] Like most accipitrid birds of prey, it builds a nest, typically in trees in this species, and is a devoted parent to a relatively small brood of young.[2] The common buzzard appears to be the most common diurnal raptor in Europe, as estimates of its total global population run well into the millions.[2][5]
Taxonomy[]
The first formal description of the common buzzard was by the Swedish naturalist Carl Linnaeus in 1758 in the tenth edition of his Systema Naturae under the binomial name Falco buteo.[6] The genus Buteo was introduced by the French naturalist Bernard Germain de Lacépède in 1799 by tautonymy with the specific name of this species.[7][8] The word buteo is Latin for a buzzard.[9] It should not be confused with the Turkey vulture, which is sometimes called a buzzard in American English.
The Buteoninae subfamily originated from and is most diversified in the Americas, with occasional broader radiations that led to common buzzards and other Eurasian and African buzzards. The common buzzard is a member of the genus Buteo, a group of medium-sized raptors with robust bodies and broad wings. The Buteo species of Eurasia and Africa are usually commonly referred to as "buzzards" while those in the Americas are called hawks. Under current classification, the genus includes approximately 28 species, the second most diverse of all extant accipitrid genera behind only Accipiter.[10] DNA testing shows that the common buzzard is fairly closely related to the red-tailed hawk (Buteo jamaicensis) of North America, which occupies a similar ecological niche to the buzzard in that continent. The two species may belong to the same species complex.[10][11] Two buzzards in Africa are likely closely related to the common buzzard based on genetic materials, the mountain (Buteo oreophilus) and forest buzzards (Buteo trizonatus), to the point where it has been questioned whether they are sufficiently distinct to qualify as full species. However, the distinctiveness of these African buzzards has generally been supported.[11][12][13] Genetic studies have further indicated that the modern buzzards of Eurasia and Africa are a relatively young group, showing that they diverged at about 300,000 years ago.[14] Nonetheless, fossils dating earlier than 5 million year old (the late Miocene period) showed Buteo species were present in Europe much earlier than that would imply, although it cannot be stated to a certainty that these would’ve been related to the extant buzzards.[15]
Subspecies and species splits[]

Some 16 subspecies have been described in the past and up to 11 are often considered valid, although some authorities accept as few as seven.[2] Common buzzard subspecies fall into two groups.
The western buteo group is mainly resident or short-distance migrants and includes:
- B. b. buteo: Ranges in Europe from the Atlantic islands, the British Isles and the Iberian Peninsula (including Madeira Island, whose population was once considered a separate race, B. b. harterti) more or less continuously throughout Europe to Finland, Romania and Asia Minor.[16] This highly individually variable race is described below. This is a relatively large and bulky race of buzzard. In males, the wing chord ranges from 350 to 418 mm (13.8 to 16.5 in) and the tail from 194 to 223 mm (7.6 to 8.8 in). In comparison, the larger female has a wing chord measuring 374 to 432 mm (14.7 to 17.0 in) and tail length of 193 to 236 mm (7.6 to 9.3 in). In both sexes, the tarsus measures 69 to 83 mm (2.7 to 3.3 in) in length.[2][4] As illustrated by average body mass, sizes in the nominate race of common buzzard seem to confirm to Bergmann's rule, increasing to the north and decreasing closer to the Equator.[17] In southern Norway, the mean weight of males was reportedly 740 g (1.63 lb), while that of females was 1,100 g (2.4 lb).[18] British buzzards were of intermediate size, 214 males averaging 781 g (1.722 lb) and 261 females averaging 969 g (2.136 lb).[17] Birds to the south in Spain were smaller, averaging 662 g (1.459 lb) in 22 males and 800 g (1.8 lb) in 30 females.[19] Cramp and Simmons (1980) listed the mean body mass overall of nominate buzzards in Europe overall as 828 g (1.825 lb) in males and 1,052 g (2.319 lb) in females.[4]

- B. b. rothschildi: This proposed race is native to the Azores islands. It is generally considered a valid subspecies.[2][4][20] This race differs from a typical intermediate of the nominate in being a darker, colder brown both above and below, closer to the darker individuals of the nominate. It averages smaller than most nominate buzzards. The wing chord of males ranges from 343 to 365 mm (13.5 to 14.4 in) while that of females ranges from 362 to 393 mm (14.3 to 15.5 in).[2][20]
- B. b. insularum: This race lives in the Canary Islands. Not all authorities consider this race suitably distinct, but others advocate it be retained as a full subspecies.[2][20] It is typically of richer brown above and more heavily streaked below compared to nominate birds. It is similar in size to B. b. rothschildi and averages slightly smaller than the nominate race. Males have a reported wing chord of 352 to 390 mm (13.9 to 15.4 in) and females have a wing chord of 370 to 394 mm (14.6 to 15.5 in).[2]
- B. b. arrigonii: This race inhabits the islands of Corsica and Sardinia. It is generally considered a valid subspecies.[2][20] The upper-side of these buzzards is an intermediate brown with very heavy streaking below, often covering the belly whereas most nominate buzzards show a whitish area the middle of the belly. Like most other insular races, this one is relatively small. Males possess a wing chord of 343 to 382 mm (13.5 to 15.0 in) while females have a wing chord of 353 to 390 mm (13.9 to 15.4 in).[2][21]

The eastern vulpinus group includes:

- B. b. vulpinus: The steppe buzzard breeds as far west as eastern Sweden, in the southern two-thirds of Finland, eastern Estonia, much of Belarus and the Ukraine, eastward to the northern Caucacus, northern Kazakhstan, Kyrgyzstan, much of Russia to Altai and south-central Siberia, Tien Shan in China and western Mongolia. B. b. vulpinus is a long-distance migrant. It winters largely in much of eastern and southern Africa. Less frequently and often very discontinuously, steppe buzzards winter in the southern peninsulas of Europe, Arabia and southwestern India in addition to some parts of southeastern Kazakhstan, Uzbekistan and Kyrgyzstan. In the open country favoured on the wintering grounds, steppe buzzards are often seen perched on roadside telephone poles.[1][2][16][22] It at one time was considered a separate species due to differences in size, form, colouring and behaviour (especially in regards to migratory behaviour) but is genetically indistinct from nominate buzzards.[4][23] Furthermore, the steppe buzzard engages in extensive interbreeding with the nominate race, mudding typical characteristics of both races. The zone of integration runs from Sweden and Finland through Eastern Europe, including any part of the overlapping ranges in the Baltic states, western Ukraine and eastern Romania. At times, the fertile hybrids of these two races have been erroneously proposed as races such as B. b. intermedius or B. b. zimmermannae. Intergrade buzzards are commonest where the grey-brown type of pale morphs of vulpinus are predominant.[2][24] Steppe buzzards are usually distinctly smaller, with relatively longer wings and tail for their size, and thus often appear swifter and more agile in flight than nominate buzzards, whose wing beats can look slower and clumsier. Typically, their length is around 45 to 50 cm (18 to 20 in), while wingspan of males average 113 cm (44 in) and females average 122.7 cm (48 in). The wing chord is 335 to 377 mm (13.2 to 14.8 in) in males and 358 to 397 mm (14.1 to 15.6 in) in females. Tail length is 170 to 200 mm (6.7 to 7.9 in) in males and 175 to 209 mm (6.9 to 8.2 in) in females.[2][16] Weights of birds from Russia can reportedly range from 560 to 675 g (1.235 to 1.488 lb) in males and 710 to 1,180 g (1.57 to 2.60 lb) in females.[25] Weights of migrant birds appear to be lower than at other times of year for steppe buzzards. Two surveys of migrant buzzards during their huge spring movement in Eilat, Israel showed 420 birds averaged 579 g (1.276 lb) and 882 birds averaged 578 g (1.274 lb).[26][27] In comparison, weights of wintering steppe buzzards was higher, averaging 725 g (1.598 lb) in 35 birds in the former Transvaal (South Africa) and 739 g (1.629 lb) in 160 birds in the Cape Province. Weights of birds from Zambia were similar.[26][28][29]
- B. b. menetriesi: This race is found in southern Crimea through the Caucasus to northern Iran and possibly into Turkey.[2][30] This race has traditionally been listed as a resident race, but some sources consider it a migrant to eastern and southern Africa.[2][31] Compared to the overlapping steppe buzzard subspecies, it is larger (roughly intermediate between the nominate race and vulpinus) and is duller in overall colour, being sandy below rather than rufous and lacking the bright rufous on the tail. Wing chord is 351 to 397 mm (13.8 to 15.6 in) in males and 372 to 413 mm (14.6 to 16.3 in) in females.[2][23]
At one time, races of the common buzzard were thought to range as far in Asia as a breeding bird well into the Himalayas and as far east as northeastern China, Russia to the Sea of Okhotsk, and all the islands of the Kurile Islands and of Japan, despite both the Himalayan and eastern birds showing a natural gap in distribution from the next nearest breeding common buzzard. However, DNA testing has revealed that the buzzards of these populations probably belong to different species. Most authorities now accept these buzzards as full species: the eastern buzzard (Buteo japonicus; with three subspecies of its own) and the Himalayan buzzard (Buteo refectus).[23][32][33] Buzzards found on the islands of Cape Verde off of the coast of western Africa, once referred to as the subspecies B. b. bannermani, and Socotra Island off of the northern peninsula of Arabia, once referred to as the rarely recognized subspecies B. b. socotrae, are now generally thought not to belong to the common buzzard. DNA testing has indicated that these insular buzzards are actually more closely related to the long-legged buzzard (Buteo rufinus) than to the common buzzard. Subsequently, some researchers have advocated full species status for the Cape Verde population, but the placement of these buzzards is generally deemed unclear.[2][20][14][34]
Description[]

The common buzzard is a medium-sized raptor that is highly variable in plumage. Most buzzards are distinctly round headed with a somewhat slender bill, relatively long wings that either reach or fall slightly short of the tail tip when perched, a fairly short tail, and somewhat short and mainly bare tarsi. They can appear fairly compact in overall appearance but may also appear large relative to other commoner raptorial birds such as kestrels and sparrowhawks.[2][35] The common buzzard measures between 40 and 58 cm (16 and 23 in) in length with a 109–140 cm (43–55 in) wingspan.[2] Females average about 2–7% larger than males linearly and weigh about 15% more. Body mass can show considerable variation. Buzzards from Great Britain alone can vary from 427 to 1,183 g (0.941 to 2.608 lb) in males, while females there can range from 486 to 1,370 g (1.071 to 3.020 lb).[2][36][37]

In Europe, most typical buzzards are dark brown above and on the upperside of the head and mantle, but can become paler and warmer brown with worn plumage. The flight feathers on perched European buzzards are always brown in the nominate subspecies (B. b. buteo). Usually the tail will usually be narrowly barred grey-brown and dark brown with a pale tip and a broad dark subterminal band but the tail in palest birds can show a varying amount a white and reduced subterminal band or even appear almost all white. In European buzzards, the underside coloring can be variable but most typically show a brown-streaked white throat with a somewhat darker chest. A pale U across breast is often present; followed by a pale line running down the belly which separates the dark areas on breast-side and flanks. These pale areas tend to have highly variable markings that tend to form irregular bars. Juvenile buzzards are quite similar to adult in the nominate race, being best told apart by having a paler eye, a narrower subterminal band on the tail and underside markings that appear as streaks rather than bars. Furthermore, juveniles may show variable creamy to rufous fringes to upperwing coverts but these also may not be present. Seen from below in flight, buzzards in Europe typically have a dark trailing edge to the wings. If seen from above, one of the best marks is their broad dark subterminal tail band. Flight feathers of typical European buzzards are largely greyish, the aforementioned dark wing linings at front with contrasting paler band along the median coverts. In flight, paler individuals tend to show dark carpal patches that can appears as blackish arches or commas but these may be indistinct in darker individuals or can appear light brownish or faded in paler individuals. Juvenile nominate buzzards are best told apart from adults in flight by the lack of a distinct subterminal band (instead showing fairly even barring throughout) and below by having less sharp and brownish rather than blackish trailing wing edge. Juvenile buzzards show streaking paler parts of under wing and body showing rather than barring as do adults.[2][35][38][39] Beyond the typical mid-range brownish buzzard, birds in Europe can range from almost uniform black-brown above to mainly white. Extreme dark individuals may range from chocolate brown to blackish with almost no pale showing but a variable, faded U on the breast and with or without faint lighter brown throat streaks. Extreme pale birds are largely whitish with variable widely spaced streaks or arrowheads of light brown about the mid-chest and flanks and may or may not show dark feather-centres on the head, wing-coverts and sometimes all but part of mantle. Individuals can show nearly endless variation of colours and hues in between these extremes and the common buzzard is counted among the most variably plumage diurnal raptors for this reason.[2][40] One study showed that this variation may actually be the result of diminished single-locus genetic diversity.[41]

Beyond the nominate form (B. b. buteo) that occupies most of the common buzzard's European range, a second main, widely distributed subspecies is known as the steppe buzzard (B. b. vulpinus). The steppe buzzard race shows three main colour morphs, each of which can be predominant in a region of breeding range. It is more distinctly polymorphic rather than just individually very variable like the nominate race. This may be because, unlike the nominate buzzard, the steppe buzzard is highly migratory. Polymorphism has been linked with migratory behaviour.[2][42] The most common type of steppe buzzard is the rufous morph which gives this subspecies its scientific name (vulpes is Latin for "fox"). This morph comprises a majority of birds seen in passage east of the Mediterranean. Rufous morph buzzards are a paler grey-brown above than most nominate B. b. buteo. Compared to the nominate race, rufous vulpinus show a patterning not dissimilar but generally far more rufous-toned on head, the fringes to mantle wing coverts and, especially, on the tail and the underside. The head is grey-brown with rufous tinges usually while the tail is rufous and can vary from almost unmarked to thinly dark-barred with a subterminal band. The underside can be uniformly pale to dark rufous, barred heavily or lightly with rufous or with dusky barring, usually with darker individuals showing the U as in nominate but with a rufous hue. The pale morph of the steppe buzzard is commonest in the west of its subspecies range, predominantly seen in winter and migration at the various land bridge of the Mediterranean. As in the rufous morph, the pale morph vulpinus is grey-brown above but the tail is generally marked with thin dark bars and a subterminal band, only showing rufous near the tip. The underside in the pale morph is greyish-white with dark grey-brown or somewhat streaked head to chest and barred belly and chest, occasionally showing darker flanks that can be somewhat rufous. Dark morph vulpinus tend to be found in the east and southeast of the subspecies range and are easily outnumbered by rufous morph while largely using similar migration points. Dark morph individuals vary from grey-brown to much darker blackish-brown, and have a tail that is dark grey or somewhat mixed grey and rufous, is distinctly marked with dark barring and has a broad, black subterminal band. Dark morph vulpinus have a head and underside that is mostly uniform dark, from dark brown to blackish-brown to almost pure black. Rufous morph juveniles are often distinctly paler in ground colour (ranging even to creamy-grey) than adults with distinct barring below actually increased in pale morph type juvenile. Pale and rufous morph juveniles can only be distinguished from each other in extreme cases. Dark morph juveniles are more similar to adult dark morph vulpinus but often show a little whitish streaking below, and like all other races have lighter coloured eyes and more evenly barred tails than adults. Steppe buzzards tend to appear smaller and more agile in flight than nominate whose wing beats can look slower and clumsier. In flight, rufous morph vulpinus have their whole body and underwing varying from uniform to patterned rufous (if patterning present, it is variable, but can be on chest and often thighs, sometimes flanks, pale band across median coverts), while the under-tail usually paler rufous than above. Whitish flight feathers are more prominent than in nominate and more marked contrast with the bold dark brown band along the trailing edges. Markings of pale vulpinus as seen in flight are similar to rufous morph (such as paler wing markings) but more greyish both on wings and body. In dark morph vulpinus the broad black trailing edges and colour of body make whitish areas of inner wing stand out further with an often bolder and blacker carpal patch than in other morphs. As in nominate, juvenile vulpinus (rufous/pale) tend to have much less distinct trailing edges, general streaking on body and along median underwing coverts. Dark morph vulpinus resemble adult in flight more so than other morphs.[2][38][39][24]
Similar species[]

The common buzzard is often confused with other raptors especially in flight or at a distance. Inexperienced and over-enthusiastic observers have even mistaken darker birds for the far larger and differently proportioned golden eagle (Aquila chrysaetos) and also dark birds for western marsh harrier (Circus aeruginosus) which also flies in a dihedral but is obviously relatively much longer and slenderer winged and tailed and with far different flying methods. Also buzzards may possibly be confused with dark or light morph booted eagles (Hieraeetus pennatus), which are similar in size, but the eagle flies on level, parallel-edged wings which usually appear broader, has a longer squarer tail, with no carpal patch in pale birds and all dark flight feathers but for whitish wedge on inner primaries in dark morph ones. Pale individuals are sometimes also mistaken with pale morph short-toed eagles (Circaetus gallicus) which are much larger with a considerably bigger head, longer wings (which are usually held evenly in flight rather than in a dihedral) and paler underwing lacking any carpal patch or dark wing lining.[2][35][38] More serious identification concerns lie in other Buteo species and in flight with honey buzzards, which are quite different looking when seen perched at close range. The European honey buzzard (Pernis apivorus) is thought in engage in mimicry of more powerful raptors, in particular, juveniles may mimic the plumage of the more powerful common buzzard.[43] While less individually variable in Europe, the honey buzzard is more extensive polymorphic on underparts than even the common buzzard. The most common morph of the adult European honey buzzard is heavily and rufous barred on the underside, quite different from the common buzzard, however the brownish juvenile much more resembles an intermediate common buzzard. Honey buzzards flap with distinctively slower and more even wing beats than common buzzard. The wings are also lifted higher on each upstroke, creating a more regular and mechanical effect, furthermore their wings are held slightly arched when soaring but not in a V. On the honey buzzard, the head appears smaller, the body thinner, the tail longer and the wings narrower and more parallel edged. The steppe buzzard race is particularly often mistaken for juvenile European honey buzzards, to the point where early observers of raptor migration in Israel considered distant individuals indistinguishable. However, when compared to a steppe buzzard, the honey buzzard has distinctly darker secondaries on the underwing with fewer and broader bars and more extensive black wing-tips (whole fingers) contrasting with a less extensively pale hand.[2][35][39] Found in the same range as the steppe buzzard in some parts of southern Siberia as well as (with wintering steppes) in southwestern India, the Oriental honey buzzard (Pernis ptilorhynchus) is larger than both the European honey buzzard and the common buzzard. The oriental species is with more similar in body plan to common buzzards, being relatively broader winged, shorter tailed and more amply-headed (though the head is still relatively small) relative to the European honey buzzard, but all plumages lack carpal patches.[2]
In much of Europe, the common buzzard is the only type of buzzard. However, the subarctic breeding rough-legged buzzard (Buteo lagopus) comes down to occupy much of the northern part of the continent during winter in the same haunts as the common buzzard. However, the rough-legged buzzard is typically larger and distinctly longer-winged with feathered legs, as well as having a white based tail with a broad subterminal band. Rough-legged buzzards have slower wing beats and hover far more frequently than do common buzzards. The carpal patch marking on the under-wing are also bolder and blacker on all paler forms of rough-legged hawk. Many pale morph rough-legged buzzards have a bold, blackish band across the belly against contrasting paler feathers, a feature which rarely appears in individual common buzzard. Usually the face also appears somewhat whitish in most pale morphs of rough-legged buzzards, which is true of only extremely pale common buzzards. Dark morph rough-legged buzzards are usually distinctly darker (ranging to almost blackish) than even extreme dark individuals of common buzzards in Europe and still have the distinct white-based tail and broad subterminal band of other roughlegs.[2][35][39] In eastern Europe and much of the Asian range of common buzzards, the long-legged buzzard (Buteo rufinus) may live alongside the common species. As in the steppe buzzard race, the long-legged buzzard has three main colour morphs that are more or less similar in hue. In both the steppe buzzard race and long-legged buzzard, the main colour is overall fairly rufous. More so than steppe buzzards, long-legged buzzards tend to have a distinctly paler head and neck compared to other feathers, and, more distinctly, a normally unbarred tail. Furthermore, the long-legged buzzard is usually a rather larger bird, often considered fairly eagle-like in appearance (although it does appear gracile and small-billed even compared to smaller true eagles), an effect enhanced by its longer tarsi, somewhat longer neck and relatively elongated wings. The flight style of the latter species is deeper, slower and more aquiline, with much more frequent hovering, showing a more protruding head and a slightly higher V held in a soar. The smaller North African and Arabian race of long-legged buzzard (B. r. cirtensis) is more similar in size and nearly all colour characteristics to steppe buzzard, extending to the heavily streaked juvenile plumage, in some cases such birds can be distinguished only by their proportions and flight patterns which remain unchanged.[2][39] Hybridization with the latter race (B. r. cirtensis) and nominate common buzzards has been observed in the Strait of Gibraltar, a few such birds have been reported potentially in the southern Mediterranean due to mutually encroaching ranges, which are blurring possibly due to climate change.[44]
Wintering steppe buzzards may live alongside mountain buzzards and especially with forest buzzard while wintering in Africa. The juveniles of steppe and forest buzzards are more or less indistinguishable and only told apart by proportions and flight style, the latter species being smaller, more compact, having a smaller bill, shorter legs and shorter and thinner wings than a steppe buzzard. However, size is not diagnostic unless side by side as the two buzzards overlap in this regard. Most reliable are the species wing proportions and their flight actions. Forest buzzard have more flexible wing beats interspersed with glides, additionally soaring on flatter wings and apparently never engage in hovering. Adult forest buzzards compared to the typical adult steppe buzzard (rufous morph) are also similar, but the forest typically has a whiter underside, sometimes mostly plain white, usually with heavy blotches or drop-shaped marks on abdomen, with barring on thighs, more narrow tear-shaped on chest and more spotted on leading edges of underwing, usually lacking marking on the white U across chest (which is otherwise similar but usually broader than that of vulpinus). In comparison, the mountain buzzard, which is more similar in size to the steppe buzzard and slightly larger than the forest buzzard, is usually duller brown above than a steppe buzzard and is more whitish below with distinctive heavy brown blotches from breasts to the belly, flanks and wing linings while juvenile mountain buzzard is buffy below with smaller and streakier markings. The steppe buzzard when compared to another African species, the red-necked buzzard (Buteo auguralis), which has red tail similar to vulpinus, is distinct in all other plumage aspects despite their similar size. The latter buzzard has a streaky rufous head and is white below with a contrasting bold dark chest in adult plumage and, in juvenile plumage, has heavy, dark blotches on the chest and flanks with pale wing-linings. Jackal and augur buzzards (Buteo rufofuscus & augur), also both rufous on the tail, are larger and bulkier than steppe buzzards and have several distinctive plumage characteristics, most notably both having their own striking, contrasting patterns of black-brown, rufous and cream.[2][35][45]
Distribution and habitat[]

The common buzzard is found throughout several islands in the eastern Atlantic islands, including the Canary Islands and Azores and almost throughout Europe. It is today found in Ireland and in nearly every part of Scotland, Wales and England. In mainland Europe, remarkably, there are no substantial gaps without breeding common buzzards from Portugal and Spain to Greece, Estonia, Belarus and Ukraine, though are present mainly only in the breeding season in much of the eastern half of the latter three countries. They are also present in all larger Mediterranean islands such as Corsica, Sardinia, Sicily and Crete. Further north in Scandinavia, they are found mainly in southeastern Norway (though also some points in southwestern Norway close to the coast and one section north of Trondheim), just over the southern half of Sweden and hugging over the Gulf of Bothnia to Finland where they live as a breeding species over nearly two-thirds of the land. The common buzzard reaches its northern limits as a breeder in far eastern Finland and over the border to European Russia, continuing as a breeder over to the narrowest straits of the White Sea and nearly to the Kola Peninsula. In these northern quarters, the common buzzard is present typically only in summer but is a year-around resident of a hearty bit of southern Sweden and some of southern Norway.[1][2][5] Outside of Europe, it is a resident of northern Turkey (largely close to the Black Sea) otherwise occurring mainly as a passage migrant or winter visitor in the remainder of Turkey, Georgia, sporadically but not rarely in Azerbaijan and Armenia, northern Iran (largely hugging the Caspian Sea) to northern Turkmenistan.[1] Further north though its absent from either side of the northern Caspian Sea, the common buzzard is found in much of western Russia (though exclusively as a breeder) including all of the Central Federal District and the Volga Federal District, all but the northernmost parts of the Northwestern and Ural Federal Districts and nearly the southern half of the Siberian Federal District, its farthest easterly occurrence as a breeder. It also found in northern Kazakhstan, Kyrgyzstan, far northwestern China (Tien Shan) and northwestern Mongolia.[1][46] Non-breeding populations occur, either as migrants or wintering birds, in southwestern India, Israel, Lebanon, Syria, Egypt (northeastern), northern Tunisia (and far northwestern Algeria), northern Morocco, near the coasts of The Gambia, Senegal and far southwestern Mauritania and Ivory Coast (and bordering Burkina Faso). In eastern and central Africa, it is found in winter from southeastern Sudan, Eritrea, about two-thirds of Ethiopia,[47] much of Kenya (though apparently absent from the northeast and northwest), Uganda, southern and eastern Democratic Republic of the Congo, and more or less the entirety of southern Africa from Angola across to Tanzania down the remainder of the continent (but for an apparent gap along the coast from southwestern Angola to northwestern South Africa).[1][48][49]
Habitat[]
The common buzzard generally inhabits the interface of woodlands and open grounds; most typically the species lives in forest edge, small woods or shelterbelts with adjacent grassland, arables or other farmland. It acquits to open moorland as long as there is some trees for perch hunting and nesting use. The woods they inhabit may be coniferous, temperate broadleaf and mixed forests and temperate deciduous forest with occasional preferences for the local dominant tree. It is absent from treeless tundra, as well as the Subarctic where the species almost entirely gives way to the rough-legged buzzard.[2][16] The common buzzard is sporadic or rare in treeless steppe but can occasionally migrate through it (despite its name, the steppe buzzard subspecies breeds primarily in the wooded fringes of the steppe).[50] The species may be found to some extent in both in mountainous or flat country. Although adaptable to and sometimes seen in wetlands and in coastal areas, buzzards are often considered more of an upland species and neither appear to be regularly attracted to or to strongly avoid bodies of waters in non-migratory times.[2][16][51] Buzzards in well-wooded areas of eastern Poland largely used large, mature stands of trees that were more humid, richer and denser than prevalent in surrounding area, but showed preference for those within 30 to 90 m (98 to 295 ft) of openings.[52] Mostly resident buzzards live in lowlands and foothills, but they can live in timbered ridges and uplands as well as rocky coasts, sometimes nesting on cliff ledges rather than trees. Buzzards may live from sea level to elevations of 2,000 m (6,600 ft), breeding mostly below 1,000 m (3,300 ft) but they can winter to an elevation of 2,500 m (8,200 ft) and migrates easily to 4,500 m (14,800 ft).[2] In the mountainous Italian Apennines, buzzard nests were at a mean elevation of 1,399 m (4,590 ft) and were, relative to the surrounding area, further from human developed areas (i.e. roads) and nearer to valley bottoms in rugged, irregularly topographed places, especially ones that faced northeast.[53] Common buzzards are fairly adaptable to agricultural lands but will show can show regional declines in apparent response to agriculture. Changes to more extensive agricultural practices were shown to reduce buzzard populations in western France where reduction of “hedgerows, woodlots and grasslands areas" caused a decline of buzzards and in Hampshire, England where more extensive grazing by free-range cattle and horses led to declines of buzzards, probably largely due to the seeming reduction of small mammal populations there.[54][55] On the contrary, buzzards in central Poland adapted to removal of pine trees and reduction of rodent prey by changing nest sites and prey for a time with no strong change in their local numbers.[56] Extensive urbanization seems to negatively affect buzzards, this species being generally less adaptable to urban areas than their New World counterparts, the red-tailed hawk. Although peri-urban areas can actually increase potential prey populations in a location at times, individual buzzard mortality, nest disturbances and nest site habitat degradation rises significantly in such areas.[57][58][59] Common buzzards are fairly adaptive to rural areas as well as suburban areas with parks and large gardens, in addition to such areas if they're near farms.[60][61][62]
Behaviour[]

The common buzzard is a typical Buteo in much of its behaviour. It is most often seen either soaring at varying heights or perched prominently on tree tops, bare branches, telegraph poles, fence posts, rocks or ledges, or alternately well inside tree canopies. Buzzards will also stand and forage on the ground. In resident populations, it may spend more than half of its day inactively perched. Furthermore, it has been described a "sluggish and not very bold" bird of prey.[16][63] It is a gifted soarer once aloft and can do so for extended periods but can appear laborious and heavy in level flight, more so nominate buzzards than steppe buzzards.[2][16] Particularly in migration, as was recorded in the case of steppe buzzards' movement over Israel, buzzards readily adjust their direction, tail and wing placement and flying height to adjust for the surrounding environment and wind conditions.[64] In Israel, migrant buzzards rarely soar all that high (maximum 1,000–2,000 m (3,300–6,600 ft) above ground) due to the lack of mountain ridges that in other areas typically produce flyways; however tail-winds are significant and allow birds to cover a mean of 9.8 metres per second (22 miles per hour).[65]
Migration[]

The common buzzard is aptly described as a partial migrant.[66] The autumn and spring movements of buzzards are subject to extensive variation, even down to the individual level, based on a region's food resources, competition (both from other buzzards and other predators), extent of human disturbance and weather conditions. Short distance movements are the norm for juveniles and some adults in autumn and winter, but more adults in central Europe and the British Isles remain on their year-around residence than do not.[2][67] Even for first year juvenile buzzards dispersal may not take them very far. In England, 96% of first-years moved in winter to less than 100 km (62 mi) from their natal site.[68] Southwestern Poland was recorded to be a fairly important wintering grounds for central European buzzards in early spring that apparently travelled from somewhat farther north, in winter average density was a locally high 2.12 individual per square kilometer.[67] Habitat and prey availability seemed to be the primary drivers of habitat selection in fall for European buzzards. In northern Germany, buzzards were recorded to show preferences in fall for areas fairly distant from nesting site, with a large quantity of vole-holes and more widely dispersed perches.[69][70] In Bulgaria, the mean wintering density was 0.34 individual per square kilometer, and buzzards showed a preference for agricultural over forested areas.[71] Similar habitat preferences were recorded in northeastern Romania, where buzzard density was 0.334–0.539 individuals per square kilometer.[72] The nominate buzzards of Scandinavia are somewhat more strongly migratory than most central European populations. However, birds from Sweden show some variation in migratory behaviours. A maximum of 41,000 individuals have been recorded at one of the main migration sites within southern Sweden in Falsterbo.[73] In southern Sweden, winter movements and migration was studied via observation of buzzard colour. White individuals were substantially more common in southern Sweden rather than further north in their Swedish range. The southern population migrates earlier than intermediate to dark buzzards, in both adults and juveniles. A larger proportion of juveniles than of adults migrate in the southern population. Especially adults in the southern population are resident to a higher degree than more northerly breeders.[74]
The entire population of the steppe buzzard is strongly migratory, covering substantial distances during migration. In no part of the range do steppe buzzards use the same summering and wintering grounds. Steppe buzzards are slightly gregarious in migration, and travel in variously sized flocks. This race migrates in September to October often from Asia Minor to the Cape of Africa in about a month but does not cross water, following around the Winam Gulf of Lake Victoria rather than crossing the several kilometer wide gulf. Similarly, they will funnel along both sides of the Black Sea. Migratory behavior of steppe buzzards mirrors those of broad-winged & Swainson's hawks (Buteo platypterus & swainsoni) in every significant way as similar long-distance migrating Buteos, including trans-equatorial movements, avoidance of large bodies of waters and flocking behaviour.[3][16] Migrating steppe buzzards will rise up with the morning thermals and can cover an average of hundreds of miles a day using the available currents along mountain ridges and other topographic features. The spring migration for steppe buzzards peaks around March–April, but the latest vulpinus arrive in their breeding grounds by late April or early May. Distances covered by migrating steppe buzzards in one way flights from northern Europe (i.e. Finland or Sweden) to southern Africa have ranged over 13,000 km (8,100 mi) within a season .[2][16] For the steppe buzzards from eastern and northern Europe and western Russia (which compromise a majority of all steppe buzzards), peak migratory numbers occur in differing areas in autumn, when the largest recorded movements occurs through Asia Minor such as Turkey, than in spring, when the largest recorded movement are to the south in the Middle East, especially Israel. The two migratory movements barely differ overall until they reach the Middle East and east Africa, where the largest volume of migrants in autumn occurs at the southern part of the Red Sea, around Djibouti and Yemen, while the main volume in spring is in the northernmost strait, around Egypt and Israel. In autumn, numbers of steppe buzzards recorded in migration have ranged up to 32,000 (recorded 1971) in northwestern Turkey (Bosporus) and in northeastern Turkey (Black Sea) up to 205,000 (recorded 1976). Further down in migration, autumn numbers of up to 98,000 have been recorded in passage in Djibouti.[2][75][76] Between 150,000 and nearly 466,000 Steppe Buzzard have been recorded migrating through Israel during spring, making this not only the most abundant migratory raptor here but one of the largest raptor migrations anywhere in the world.[65][77] Migratory movements of southern Africa buzzards largely occur along the major mountain ranges, such as the Drakensberg and Lebombo Mountains.[45] Wintering steppe buzzards occur far more irregularly in Transvaal than Cape region in winter.[78] The onset of migratory movement for steppe buzzards back to the breeding grounds in southern Africa is mainly in March, peaking in the second week.[45] Steppe buzzard molt their feathers rapidly upon arrival at wintering grounds and seems to split their flight feather molt between breeding ground in Eurasia and wintering ground in southern Africa, the molt pausing during migration.[79] In last 50 years, it was recorded that nominate buzzards are typically migrating shorter distances and wintering further north, possibly in response to climate change, resulting in relatively smaller numbers of them at migration sites. They are also extending their breeding range possibly reducing/supplanting steppe buzzards.[74][80][81]
Vocalizations[]
Resident populations of common buzzards tend to vocalize all year around, whereas migrants tend to vocalize only during the breeding season. Both nominate buzzards and steppe buzzards (and their numerous related subspecies within their types) tend to have similar voices. The main call of the species is a plaintive, far-carrying pee-yow or peee-oo, used as both contact call and more excitedly in aerial displays. Their call is sharper, more ringing when used in aggression, tends to be more drawn-out and wavering when chasing intruders, sharper, more yelping when as warning when approaching the nest or shorter and more explosive when called in alarm. Other variations of their vocal performances include a cat-like mew, uttered repeatedly on the wing or when perched, especially in display; a repeated mah has been recorded as uttered by pairs answering each other, further chuckles and croaks have also been recorded at nests.[2][16][82] Juveniles can usually be distinguished by the discordant nature of their calls compared to those of adults.[83]
Dietary biology[]

The common buzzard is a generalist predator which hunts a wide variety of prey given the opportunity. Their prey spectrum extents to a wide variety of vertebrates including mammals, birds (from any age from eggs to adult birds), reptiles, amphibians and, rarely, fish, as well as to various invertebrates, mostly insects. Young animals are often attacked, largely the nidifugous young of various vertebrates. In total well over 300 prey species are known to be taken by common buzzards. Furthermore, prey size can vary from tiny beetles, caterpillars and ants to large adult grouse and rabbits up to nearly twice their body mass.[2][16][84][85][86] Mean body mass of vertebrate prey was estimated at 179.6 g (6.34 oz) in Belarus.[87] At times, they will also subsist partially on carrion, usually of dead mammals or fish.[2] However, dietary studies have shown that they mostly prey upon small mammals, largely small rodents.[47] Like many temperate zone raptorial birds of varied lineages, voles are an essential part of the common buzzard's diet. This bird's preference for the interface between woods and open areas frequently puts them in ideal vole habitat.[88][89] Hunting in relatively open areas has been found to increase hunting success whereas more complete shrub cover lowered success.[90] A majority of prey is taken by dropping from perch, and is normally taken on ground. Alternately, prey may be hunted in a low flight. This species tends not to hunt in a spectacular stoop but generally drops gently then gradually accelerate at bottom with wings held above the back. Sometimes, the buzzard also forages by random glides or soars over open country, wood edges or clearings. Perch hunting may be done preferentially but buzzards fairly regularly also hunt from a ground position when the habitat demands it. Outside the breeding season, as many 15–30 buzzards have been recorded foraging on ground in a single large field, especially juveniles. Normally the rarest foraging type is hovering. A study from Great Britain indicated that hovering does not seem to increase hunting success.[2][16][91]
Mammals[]

A high diversity of rodents may be taken given the chance, as around 60 species of rodent have been recorded in the foods of common buzzards.[84][85] It seems clear that voles are the most significant prey type for European buzzards. Nearly every study from the continent makes reference to the importance, in particular, of the two most numerous and widely distributed European voles: the 28.5 g (1.01 oz) common vole (Microtus arvalis) and the somewhat more northerly ranging 40 g (1.4 oz) field vole (Microtus agrestis).[92][93][94] In southern Scotland, field voles were the best represented species in pellets, accounting for 32.1% of 581 pellets.[95][96] In southern Norway, field voles were again the main food in years with peak vole numbers, accounting for 40.8% of 179 prey items in 1985 and 24.7% of 332 prey items in 1994. Altogether, rodents amount to 67.6% and 58.4% of the foods in these respective peak vole years. However, in low vole population years, the contribution of rodents to the diet was minor.[93] As far west as the Netherlands, common voles were the most regular prey, amounting to 19.6% of 6624 prey items in a very large study.[97] Common voles were the main foods recorded in central Slovakia, accounting for 26.5% of 606 prey items.[98] The common vole, or other related vole species at times, were the main foods as well in the Ukraine (17.2% of 146 prey items) ranging east to Russia in the Privolshky Steppe Nature Reserve (41.8% of 74 prey items) and in Samara (21.4% of 183 prey items).[84] Other records from Russia and the Ukraine show voles ranging from slightly secondary prey to as much as 42.2% of the diet.[99] In Belarus, voles, including Microtus species and 18.4 g (0.65 oz) bank voles (Myodes glareolus), accounted for 34.8% of the biomass on average in 1065 prey items from different study areas over 4 years.[87] At least 12 species of the genus Microtus are known to be hunted by common buzzards and even this is probably conservative, moreover similar species like lemmings will be taken if available.[84][86][98][99][100]
Other rodents are taken largely opportunistically rather than by preference. Several wood mice (Apodemus ssp.) are known to be taken quite frequently but given their preference for activity in deeper woods than the field-forest interfaces preferred, they are rarely more than secondary food items.[92][93] An exception was in Samara where the yellow-necked mouse (Apodemus flavicollis), one of the largest of its genus at 28.4 g (1.00 oz), made up 20.9%, putting it just behind the common vole in importance.[84] Similarly, tree squirrels are readily taken but rarely important in the foods of buzzards in Europe, as buzzards apparently prefer to avoid taking prey from trees nor do they possess the agility typically necessary to capture significant quantities of tree squirrels.[16][85][101][102] All four ground squirrels that range (mostly) into eastern Europe are also known to be common buzzard prey but little quantitative analysis has gone into how significant such predator-prey relations are.[103][104][105][106] Rodent prey taken have ranged in size from the 7.8 g (0.28 oz) Eurasian harvest mouse (Micromys minutus) to the non-native, 1,100 g (2.4 lb) muskrat (Ondatra zibethicus).[107] Other rodents taken either seldomly or in areas where the food habits of buzzards are spottily known include flying squirrels, marmots (presumably very young if taken alive), chipmunks, spiny rats, hamsters, mole-rats, gerbils, jirds and jerboas and occasionally hearty numbers of dormice, although these are nocturnal.[79][87][98][99][108][109][110][111][112] Surprisingly little research has gone into the diets of wintering steppe buzzards in southern Africa, considering their numerous status there. However, it has been indicated that the main prey remains consist of rodents such as the four-striped grass mouse (Rhabdomys pumilio) and Cape mole-rats (Georychus capensis).[45][113]
Other than rodents, two other groups of mammals can be counted as significant to the diet of common buzzards. One of these main prey type of import in the diets of common buzzards are leporids or lagomorphs, especially the European rabbit (Oryctolagus cuniculus) where it is found in numbers in a wild or feral state. In all dietary studies from Scotland, rabbits were highly important to the buzzard's diet. In southern Scotland, rabbits constituted 40.8% of remains at nests and 21.6% of pellet contents, while lagomorphs (mainly rabbits but also some young hares) were present in 99% of remains in Moray, Scotland. The nutritional richness relative to the commonest prey elsewhere, such as voles, might account for the high productivity of buzzards here. For example, clutch sizes were twice as large on average where rabbits were common (Moray) than were where they were rare (Glen Urquhart).[95][114][115] In northern Ireland, an area of interest because it is devoid of any native vole species, rabbits were again the main prey. Here, lagomorphs constituted 22.5% of prey items by number and 43.7% by biomass.[116] While rabbits are non-native, albeit long-established, in the British Isles, in their native area of the Iberian peninsula, rabbits are similarly significant to the buzzard's diet. In Murcia, Spain, rabbits were the most common mammal in the diet, making up 16.8% of 167 prey items.[86] In a large study from northeastern Spain, rabbits were dominant in the buzzard's foods, making up 66.5% of 598 prey items.[117] In the Netherlands, European rabbits were second in number (19.1% of 6624 prey items) only to common voles and the largest contributor of biomass to nests (36.7%).[97] Outside of these (at least historically) rabbit-rich areas, leverets of the common hare species found in Europe can be important supplemental prey. European hare (Lepus europaeus) were the fourth most important prey species in central Poland and the third most significant prey species in Stavropol Krai, Russia.[99][110] Buzzards normally attack the young of European rabbits, which as adults can average nearly 2,000 g (4.4 lb), and invariably (so far as is known) only the young of hares, which can average up to twice as massive as rabbits. The mean weights of rabbits taken have various been estimated from 159 to 550 g (5.6 to 19.4 oz) in different areas while mountain hares (Lepus timidus) taken in Norway were estimated to average about 1,000 g (2.2 lb), in both cases about a third of the weight of full-grown, prime adults of the respective species. However, hares and rabbits taken by female buzzards can infrequently include specimens that weigh up to 1,600 g (3.5 lb), including at times adult rabbits.[85][97][95][116][114][118][119]
The other significant mammalian prey type is insectivores, among which more than 20 species are known to be taken by this species, including nearly all the species of shrew, mole and hedgehog found in Europe. Moles are taken particularly often among this order, since as is the case with "vole-holes", buzzard probably tend to watch molehills in fields for activity and dive quickly from their perch when one of the subterranean mammals pops up.[120] The most widely found mole in the buzzard's northern range is the 98 g (3.5 oz) European mole (Talpa europaea) and this is one of the more important non-rodent prey items for the species. This species was present in 55% of 101 remains in Glen Urquhart, Scotland and was the second most common prey species (18.6%) in 606 prey items in Slovakia.[98][114] In Bari, Italy, the Roman mole (Talpa romana), of similar size to the European species, was the leading identified mammalian prey, making up 10.7% of the diet.[121] The full size range of insectivores may be taken by buzzards, ranging from the world's smallest mammal (by weight), the 1.8 g (0.063 oz) Etruscan shrew (Suncus etruscus) to arguably the heaviest insectivore, the 800 g (28 oz) European hedgehog (Erinaceus europaeus).[114][117] Mammalian prey for common buzzards other than rodents, insectivores and lagomorphs is rarely taken. Occasionally, some weasels (including polecats) and perhaps martens might be attacked by buzzards, more likely the more powerful female buzzard since such prey is potentially dangerous and of similar size to a buzzard itself.[93][97][99][95][122] Numerous larger mammals, including medium-sized carnivores such as dogs, cats and foxes and various ungulates, are sometimes eaten as carrion by buzzards, mainly during lean winter months. Still-borns of deer are also visited with some frequency.[87][114][106][123]
Birds[]

When attacking birds, common buzzards chiefly prey on nestlings and fledglings of small to medium-sized birds, largely passerines but also a variety of gamebirds, but sometimes also injured, sickly or unwary but healthy adults. While capable of overpowering birds larger than itself, the common buzzard is usually considered to lack the agility necessary to capture many adult birds, even gamebirds which would presumably be weaker fliers considering their relatively heavy bodies and small wings.[2][16] The amount of fledgling and younger birds preyed upon relative to adults is variable, however. For example, in the Italian Alps, 72% of birds taken were fledglings or recently fledged juveniles, 19% were nestlings and 8% were adults.[124] On the contrary, in southern Scotland, even though the buzzards were taking relatively large bird prey, largely red grouse (Lagopus lagopus scotica), 87% of birds taken were reportedly adults.[95] In total, as in many raptorial birds that are far from bird-hunting specialists, birds are the most diverse group in the buzzard's prey spectrum due to the sheer number and diversity of birds, few raptors do not hunt them at least occasionally. Nearly 150 species of bird have been identified in the common buzzard's diet. In general, despite many that are taken, birds usually take a secondary position in the diet after mammals.[93][97][99] In northern Scotland, birds were fairly numerous in the foods of buzzards. The most often recorded avian prey and 2nd and 3rd most frequent prey species (after only field voles) in Glen Urquhart, were 23.9 g (0.84 oz) chaffinch (Fringilla coelebs) and 18.4 g (0.65 oz) meadow pipits (Anthus pratensis), with the buzzards taking 195 fledglings of these species against only 90 adults. This differed from Moray where the most frequent avian prey and 2nd most frequent prey species behind the rabbit was the 480 g (17 oz) common wood pigeon (Columba palumbus) and the buzzards took four times as many adults relative to fledglings.[114]
Birds were the primary food for common buzzards in the Italian Alps, where they made up 46% of the diet against mammal which accounted for 29% in 146 prey items. The leading prey species here were 103 g (3.6 oz) Eurasian blackbirds (Turdus merula) and 160 g (5.6 oz) Eurasian jays (Garrulus glandarius), albeit largely fledglings were taken of both.[124] Birds could also take the leading position in years with low vole populations in southern Norway, in particular thrushes, namely the blackbird, the 67.7 g (2.39 oz) song thrush (Turdus philomelos) and the 61 g (2.2 oz) redwing (Turdus iliacus), which were collectively 22.1% of 244 prey items in 1993.[93] In southern Spain, birds were equal in number to mammals in the diet, both at 38.3%, but most remains were classified as "unidentified medium-sized birds", although the most often identified species of those that apparently could be determined were Eurasian jays and red-legged partridges (Alectoris rufa).[86] Similarly, in northern Ireland, birds were roughly equal in import to mammals but most were unidentified corvids.[116] In Seversky Donets, Ukraine, birds and mammals both made up 39.3% of the foods of buzzards.[99] Common buzzards may hunt nearly 80 species passerines and nearly all available gamebirds. Like many other largish raptors, gamebirds are attractive to hunt for buzzards due to their ground-dwelling habits. Buzzards were the most frequent predator in a study of juvenile pheasants in England, accounting for 4.3% of 725 deaths (against 3.2% by foxes, 0.7% by owls and 0.5% by other mammals).[90][125] They also prey on a wide size range of birds, ranging down to Europe's smallest bird, the 5.2 g (0.18 oz) goldcrest (Regulus regulus).[17][93] Very few individual birds hunted by buzzards weigh more than 500 g (1.1 lb). However, there have been some particularly large avian kills by buzzards, including any that weigh more or 1,000 g (2.2 lb), or about the largest average size of a buzzard, have including adults of mallard (Anas platyrhynchos), black grouse (Tetrao tetrix), ring-necked pheasant (Phasianus colchicus), common raven (Corvus corax) and some of the larger gulls if ambushed on their nests. The largest avian kill by a buzzard, and possibly largest known overall for the species, was an adult female western capercaillie (Tetrao urogallus) that weighed an estimated 1,985 g (4.376 lb).[93][95][97][98][114] At times, buzzards will hunt the young of large birds such as herons and cranes.[126][127] Other assorted avian prey has included a few species of waterfowl, most available pigeons and doves, cuckoos, swifts, grebes, rails, nearly 20 assorted shorebirds, tubenoses, hoopoes, bee-eaters and several types of woodpecker. Birds with more conspicuous or open nesting areas or habits are more likely to have fledglings or nestlings attacked, such as water birds, while those with more secluded or inaccessible nests, such as pigeons/doves and woodpeckers, adults are more likely to be hunted.[86][97][98][106][114][128][129][130]
Reptiles and amphibians[]

The common buzzard may be the most regular avian predator of reptiles and amphibians in Europe apart from the sections where they are sympatric with the largely snake-eating short-toed eagle.[131] In total, the prey spectrum of common buzzards include nearly 50 herpetological prey species. In studies from northern and southern Spain, the leading prey numerically were both reptilian, although in Biscay (northern Spain) the leading prey (19%) was classified as "unidentified snakes". In Murcia, the most numerous prey was the 77.2 g (2.72 oz) ocellated lizard (Timon lepidus), at 32.9%. In total, at Biscay and Murcia, reptiles accounted for 30.4% and 35.9% of the prey items, respectively. Findings were similar in a separate study from northeastern Spain, where reptiles amounted to 35.9% of prey.[86][117][132] In Bari, Italy, reptiles were the main prey, making up almost exactly half of the biomass, led by the large green whip snake (Hierophis viridiflavus), maximum size up to 1,360 g (3.00 lb), at 24.2% of food mass.[121] In Stavropol Krai, Russia, the 20 g (0.71 oz) sand lizard (Lacerta agilis) was the main prey at 23.7% of 55 prey items.[99] The 16 g (0.56 oz) slowworm (Anguis fragilis), a legless lizard, became the most numerous prey for the buzzards of southern Norway in low vole years, amounting to 21.3% of 244 prey items in 1993 and were also common even in the peak vole year of 1994 (19% of 332 prey items).[93] More or less any snake in Europe is potential prey and the buzzard has been known to be uncharacteristically bold in going after and overpowering large snakes such as rat snakes, ranging up to nearly 1.5 m (4 ft 11 in) in length, and healthy, large vipers despite the danger of being struck by such prey.[93][133][134][135] However, in at least one case, the corpse of a female buzzard was found envenomed over the body of an adder that it had killed.[136] In some parts of range, the common buzzard acquires the habit of taking many frogs and toads.[2] This was the case in the Mogilev Region of Belarus where the 23 g (0.81 oz) moor frog (Rana arvalis) was the major prey (28.5%) over several years, followed by other frogs and toads amounting to 39.4% of the diet over the years.[99] In central Scotland, the 46 g (1.6 oz) common toad (Bufo bufo) was the most numerous prey species, accounting for 21.7% of 263 prey items, while the common frog (Rana temporaria) made up a further 14.7% of the diet.[115] Frogs made up about 10% of the diet in central Poland as well.[110]
Invertebrates and other prey[]
When common buzzards feed on invertebrates, these are chiefly earthworms, beetles and caterpillars in Europe and largely seemed to be preyed on by juvenile buzzards with less refined hunting skills or in areas with mild winters and ample swarming or social insects. In most dietary studies, invertebrates are at best a minor supplemental contributor to the buzzard's diet.[4][98] Nonetheless, roughly a dozen beetle species have found in the foods of buzzards from the Ukraine alone.[84] In winter in northeastern Spain, it was found that the buzzards switched largely from the vertebrate prey typically taken during spring and summer to a largely insect-based diet. Most of this prey was unidentified but the most frequently identified were European mantis (Mantis religiosa) and European mole cricket (Gryllotalpa gryllotalpa).[117] In the Ukraine, 30.8% of the food by number was found to be insects.[84] Especially in winter quarters such as southern Africa, common buzzards are often attracted to swarming locusts and other orthopterans.[2][45] In this way the steppe buzzard may mirror a similar long-distance migrant from the Americas, the Swainson's hawk, which feeds its young largely on nutritious vertebrates but switches to a largely insect-based once the reach their distant wintering grounds in South America.[3] In , 18 returning migrant steppe buzzards were seen to feed together on swarms of grasshoppers.[137] For wintering steppe buzzards in Zimbabwe, one source went so far as to refer to them as primarily insectivorous, apparently being somewhat locally specialized to feeding on termites.[138] Stomach contents in buzzards from Malawi apparently consisted largely of grasshoppers (alternately with lizards).[139] Fish tend to be the rarest class of prey found in the common buzzard's foods. There are a couple cases of predation of fish detected in the Netherlands, while elsewhere they've been known to have fed upon eels and carp.[97][140][141]
Interspecies predatory relationships[]

Common buzzards co-occur with dozens of other raptorial birds through their breeding, resident and wintering grounds. There may be many other birds that broadly overlap in prey selection to some extent. Furthermore, their preference for interferences of forest and field is used heavily by many birds of prey. Some of the most similar species by diet are the common kestrel (Falco tinniculus), hen harrier (Circus cyaenus) and lesser spotted eagle (Clanga clanga), not to mention nearly every European species of owl, as all but two may locally prefer rodents such as voles in their diets.[142][143][144][145][146] Diet overlap was found to be extensive between buzzards and red foxes (Vulpes vulpes) in Poland, with 61.9% of prey selection overlapping by species although the dietary breadth of the fox was broader and more opportunistic. Both fox dens and buzzard roosts were found to be significantly closer to high vole areas relative to the overall environment here.[147] The only other widely found European Buteo, the rough-legged buzzard, comes to winter extensively with common buzzards. It was found in southern Sweden, habitat, hunting and prey selection often overlapped considerably. Rough-legged buzzards appear to prefer slightly more open habitat and took slightly fewer wood mice than common buzzard. Roughlegs also hover much more frequently and are more given to hunting in high winds. The two buzzards are aggressive towards one another and excluded each other from winter feeding territories in similar ways to the way they exclude conspecifics. In northern Germany, the buffer of their habitat preferences apparently accounted for the lack of effect on each other's occupancy between the two buzzard species.[148][69] Despite a broad range of overlap, very little is known about the ecology of common and long-legged buzzards where they co-exist. However, it can be inferred from the long-legged species preference for predation on differing prey, such as blind mole-rats, ground squirrels, hamsters and gerbils, from the voles usually preferred by the common species, that serious competition for food is unlikely.[149][150]
A more direct negative effect has been found in buzzard's co-existence with northern goshawk (Accipiter gentilis). Despite the considerable discrepancy of the two species dietary habits, habitat selection in Europe is largely similar between buzzards and goshawks. Goshawks are slightly larger than buzzards and are more powerful, agile and generally more aggressive birds, and so they are considered dominant. In studies from Germany and Sweden, buzzards were found to be less disturbance sensitive than goshawks but were probably displaced into inferior nesting spots by the dominant goshawks. The exposure of buzzards to a dummy goshawk was found to decrease breeding success whereas there was no effect on breeding goshawks when they were exposed to a dummy buzzard.[151][152][153] In many cases, in Germany and Sweden, goshawks displaced buzzards from their nests to take them over for themselves.[154][155] In Poland, buzzards productivity was correlated to prey population variations, particularly voles which could vary from 10–80 per hectare, whereas goshawks were seemingly unaffected by prey variations; buzzards were found here to number 1.73 pair per 10 km2 (3.9 sq mi) against goshawk 1.63 pair per 10 km2 (3.9 sq mi).[156] In contrast, the slightly larger counterpart of buzzards in North America, the red-tailed hawk (which is also slightly larger than American goshawks, the latter averaging smaller than European ones) are more similar in diet to goshawks there. Redtails are not invariably dominated by goshawks and are frequently able to outcompete them by virtue of greater dietary and habitat flexibility. Furthermore, red-tailed hawks are apparently equally capable of killing goshawks as goshawks are of killing them (killings are more one-sided in buzzard-goshawk interactions in favour of the latter).[157][158][159] Other raptorial birds, including many of similar or mildly larger size than common buzzards themselves, may dominate or displace the buzzard, especially with aims to take over their nests. Species such as the black kite (Milvus migrans), booted eagle (Hieraeetus pennatus) and the lesser spotted eagle have been known to displace actively nesting buzzards, although in some cases the buzzards may attempt to defend themselves. The broad range of accipitrids that take over buzzard nests is somewhat unusual. More typically, common buzzards are victims of nest parasitism to owls and falcons, as neither of these other kinds of raptorial birds builds their own nests, but these may regularly take up occupancy on already abandoned or alternate nests rather than ones the buzzards are actively using.[124][100][160][161] Even with birds not traditionally considered raptorial, such as common ravens, may compete for nesting sites with buzzards.[162] In urban vicinities of southwestern England, it was found that peregrine falcons (Falco peregrinus) were harassing buzzards so persistently, in many cases resulting in injury or death for the buzzards, the attacks tending to peak during the falcon's breeding seasons and tend to be focused on subadult buzzards.[163] Despite often being dominated in nesting site confrontations by even similarly sized raptors, buzzards appear to be bolder in direct competition over food with other raptors outside of the context of breeding, and has even been known to displace larger birds of prey such as red kites (Milvus milvus) and female buzzards may also dominate male goshawks (which are much smaller than the female goshawk) at disputed kills.[155][164]

Common buzzards are occasionally threatened by predation by other raptorial birds. Northern goshawks have been known to have preyed upon buzzards in a few cases.[165][166] Much larger raptors are known to have killed a few buzzards as well, including steppe eagles (Aquila nipalensis) on migrating steppe buzzards in Israel. Further instances of predation on buzzards have involved golden, eastern imperial (Aquila heliaca), Bonelli's (Aquila fasciata) and white-tailed eagles (Haliaeetus albicilla) in Europe.[167][168][169][170][171][172] Besides preying on adult buzzard, white-tailed eagles have been known to raise buzzards with their own young. These are most likely cases of eagles carrying off young buzzard nestlings with the intention of predation but, for unclear reasons, not killing them. Instead the mother eagle comes to brood the young buzzard. Despite the difference of the two species diets, white-tailed eagles are surprisingly successful at raising young buzzards (which are conspicuously much smaller than their own nestlings) to fledging.[173][174] Studies in Lithuania of white-tailed eagle diets found that predation on common buzzards was more frequent than anticipated, with 36 buzzard remains found in 11 years of study of the summer diet of the white-tailed eagles. While nestling buzzards were multiple times more vulnerable to predation than adult buzzards in the Lithuanian data, the region's buzzards expelled considerable time and energy during the late nesting period trying to protect their nests.[175][176] The most serious predator of common buzzards, however, is almost certainly the Eurasian eagle-owl (Bubo bubo). This is a very large owl with a mean body mass about three to four times greater than that of a buzzard. The eagle-owl, despite often taking small mammals that broadly overlap with those selected by buzzards, is considered a "super-predator" that is a major threat to nearly all co-existing raptorial birds, capably destroying whole broods of other raptorial birds and dispatching adult raptors even as large as eagles. Due to their large numbers in edge habitats, common buzzards frequently feature heavily in the eagle-owl's diet. Eagle-owls, as will some other large owls, also readily expropriate the nests of buzzards.[165][177][178] In the Czech Republic and in Luxembourg, the buzzard was the third and fifth most frequent prey species for eagle-owls, respectively.[179][180] The reintroduction of eagle-owls to sections of Germany has been found to have a slight deleterious effect on the local occupancy of common buzzards. The only sparing factor is the temporal difference (the buzzard nesting later in the year than the eagle-owl) and buzzards may locally be able to avoid nesting near an active eagle-owl family.[181] As the ecology of the wintering population is relatively little studied, a similar very large owl at the top of the avian food chain, the Verreaux's eagle-owl (Bubo lacteus), is the only known predator of wintering steppe buzzards in southern Africa.[182] Despite not being known predators of buzzards, other large, vole-eating owls are known to displace or to be avoided by nesting buzzards, such as great grey owls (Strix nebulosa) and Ural owls (Strix uralensis).[166][183] Unlike with large birds of prey, next to nothing is known of mammalian predators of common buzzards, despite up to several nestlings and fledglings being likely depredated by mammals.[16][184][185]
Common buzzards themselves rarely present a threat to other raptorial birds but may occasionally kill a few of those of smaller size. The buzzard is a known predator of 237 g (8.4 oz) Eurasian sparrowhawks (Accipiter nisus), 184 g (6.5 oz) common kestrel and 152 g (5.4 oz) lesser kestrel (Falco naumanni) .[107][186] Perhaps surprisingly, given the nocturnal habits of this prey, the group of raptorial birds the buzzard is known to hunt most extensively is owls.[187] Known owl prey has included 419 g (14.8 oz) barn owls (Tyto alba), 92 g (3.2 oz) European scops owls (Otus scops), 475 g (16.8 oz) tawny owls (Strix alucco), 169 g (6.0 oz) little owls (Athene noctua), 138 g (4.9 oz) boreal owls (Aegolius funereus), 286 g (10.1 oz) long-eared owls (Asio otus) and 355 g (12.5 oz) short-eared owls (Asio flammeus).[98][117][114][107][188][189] Despite their relatively large size, tawny owls are known to avoid buzzards as there are several records of them preying upon the owls.[97][190][191]
Breeding[]

Nesting territories and density[]
Home ranges of common buzzards are generally 0.5 to 2 km2 (0.19 to 0.77 sq mi).[16] The size of breeding territory seem to be generally correlated with food supply.[16][192] In a German study, the range was 0.8 to 1.8 km2 (0.31 to 0.69 sq mi) with an average of 1.26 km2 (0.49 sq mi).[69] Some of the lowest pair densities of common buzzards seem to come from Russia. For instance, in Kerzhenets Nature Reserve, the recorded density was 0.6 pairs per 100 km2 (39 sq mi) and the average distance of nearest neighbors was 3.8 km (2.4 mi).[193] The Snowdonia region of northern Wales held a pair per 9.7 km2 (3.7 sq mi) with a mean nearest neighbor distance of 1.95 km (1.21 mi); in adjacent Migneint, pair occurrence was 7.2 km2 (2.8 sq mi), with a mean distance of 1.53 km (0.95 mi).[194] In the Teno massif of the Canary Islands, the average density was estimated as 23 pairs per 100 km2 (39 sq mi), similar to that of a middling continental population.[195] On another set of islands, on Crete the density of pairs was lower at 5.7 pairs per 100 km2 (39 sq mi); here buzzards tend to have an irregular distribution, some in lower intensity harvest olive groves but their occurrence actually more common in agricultural than natural areas.[196] In the Italian Alps, it was recorded in 1993–96 that there were from 28 to 30 pairs per 100 km2 (39 sq mi).[124] In central Italy, density average was lower at 19.74 pairs per 100 km2 (39 sq mi).[197] Higher density areas are known than those above. Two areas of the Midlands of England showed occupancies of 81 and 22 territorial pairs per 100 km2 (39 sq mi). High buzzard densities there were associated with high proportions of unimproved pasture and mature woodland within the estimated territories.[91] Similarly high densities of common buzzards were estimated in central Slovakia using two different methods, here indicating densities of 96 to 129 pairs per 100 km2 (39 sq mi).[198] Despite claims from the study of the English midlands were the highest known territory density for the species, a number ranging from 32 to 51 pairs in wooded area of merely 22 km2 (8.5 sq mi) in Czech Republic seems to surely exceed even those densities. The Czech study hypothesized that fragmentation of forest in human management of lands for wild sheep and deer, creating exceptional concentrations of prey such as voles, and lack of appropriate habitat in surrounding regions for the exceptionally high density.[91][199]

Common buzzards maintain their territories through flight displays. In Europe, territorial behaviour generally starts in February. However, displays are not uncommon throughout year in resident pairs, especially by males, and can elicit similar displays by neighbors.[2][16] In them, common buzzards generally engage in high circling, spiraling upward on slightly raised wings. Mutual high circling by pairs sometimes go on at length, especially during the period prior to or during breeding season. In mutual displays, a pair may follow each other at 10–50 m (33–164 ft) in level flight. During the mutual displays, the male may engage in exaggerated deep flapping or zig-zag tumbling, apparently in response to the female being too distant. Two or three pairs may circle together at times and as many as 14 individual adults have been recorded over established display sites.[2][200] Sky-dancing by common buzzards have been recorded in spring and autumn, typically by male but sometimes by female, nearly always with much calling. Their sky-dances are of the rollercoaster type, with upward sweep until they start to stall, but sometimes embellished with loops or rolls at the top. Next in the sky-dance, they dive on more or less closed wings before spreading them and shooting up again, upward sweeps of up to 30 m (98 ft), with dive drops of up to at least 60 m (200 ft). These dances may be repeated in series of 10 to 20. In the climax of the sky dance, the undulations become progressive shallower, often slowing and terminating directly onto a perch. Various other aerial displays include low contour flight or weaving among trees, frequently with deep beats and exaggerated upstrokes which show underwing pattern to rivals perched below.[2][201] Talon grappling and occasionally cartwheeling downward with feet interlocked has been recorded in buzzards and, as in many raptors, is likely the physical culmination of the aggressive territorial display, especially between males.[16][202] Despite the highly territorial nature of buzzards and their devotion to a single mate and breeding ground each summer, there is one case of a polyandrous trio of buzzards nesting in the Canary Islands.[203] In North-Estonian Neeruti landscape reserve (area 1272 ha) found in years 1989 and 1990 Marek Vahula 9 populated nest. This is sovereign public density of population. One nest founded in 12.06.1982 and this is apparently oldest nest of Common Buzzard, what is populated until today.
Nests[]
Common buzzards tend to build a bulky nest of sticks, twigs and often heather. Commonly, nests are up to 1 to 1.2 m (3 ft 3 in to 3 ft 11 in) across and 60 cm (24 in) deep. With reuse over years, the diameter can reach or exceed 1.5 m (4 ft 11 in) and weight of nests can reach over 200 kg (440 lb).[2][16] Active nests tend to be lined with greenery, most often this consists of broad-leafed foliage but sometimes also includes rush or seaweed locally. Nest height in trees is commonly 3 to 25 m (9.8 to 82.0 ft), usually by main trunk or main crutch of the tree.[2][16][4] In Germany, trees used for nesting consisted mostly of red beeches (Fagus sylvatica) (in 337 cases), whereas a further 84 were in assorted oaks.[204] Buzzards were recorded to nest almost exclusively in pines in Spain at a mean height of 14.5 m (48 ft).[86] Trees are generally used for a nesting location but they will also utilize crags or bluffs if trees are unavailable.[16] Buzzards in one English study were surprisingly partial to nesting on well-vegetated banks and due to the rich surrounding environment habitat and prey population, were actually more productive than nests located in other locations here.[205] Furthermore, a few ground nests were recorded in high prey-level agricultural areas in the Netherlands.[206] In the Italian Alps, 81% of 108 nests were on cliffs.[124] The common buzzard generally lacks the propensity of its Nearctic counterpart, the red-tailed hawk, to occasionally nest on or near manmade structures (often in heavily urbanized areas) but in Spain some pairs recorded nesting along the perimeter of abandoned buildings.[207] Pairs often have several nests but some pairs may use one over several consecutive years. Two to four alternate nests in a territory is typical for common buzzards, especially those breeding further north in their range.[16][208]
Reproduction and eggs[]

The breeding season commences at differing times based on latitude. Common buzzard breeding seasons may fall as early as January to April but typically the breeding season is March to July in much of Palearctic. In the northern stretches of the range the breeding season may last into May–August. Mating usually occurs on or near the nest and lasts about 15 seconds, typically occurring several times a day.[2][16] Eggs are usually laid in 2 to 3-day intervals. The clutch size can range from to 2 to 6, a relatively large clutch for an accipitrid. More northerly and westerly buzzard usually bear larger clutches, which average nearer 3, than those further east and south.[2][16][195] In Spain, the average clutch size is about 2 to 2.3.[86] From 4 locations in different parts of Europe, 43% had clutch size of 2, 41% had size of 3, clutches of 1 and 4 each constituted about 8%.[209] Laying dates are remarkably constant throughout Great Britain. There are, however, highly significant differences in clutch size between British study areas. These do not follow any latitudinal gradient and it is likely that local factors such as habitat and prey availability are more important determinants of clutch size.[184] The eggs are white in ground colour, rather round in shape with sporadic red to brown markings sometimes lightly showing. In the nominate race, egg size is 49.8–63.8 mm (1.96–2.51 in) in height by 39.1–48.2 mm (1.54–1.90 in) in diameter with an average of 55 mm × 44 mm (2.2 in × 1.7 in) in 600 eggs. In the race of vulpinus, egg height is 48–63 mm (1.9–2.5 in) by 39.2–47.5 mm (1.54–1.87 in) with an average of 54.2 mm × 42.8 mm (2.13 in × 1.69 in) in 303 eggs.[16] Eggs are generally laid in late March to early April in extreme south, sometime in April in most of Europe, into May and possibly even early June in the extreme north. If eggs are lost to a predator (including humans) or fail in some other way, common buzzards do not usually lay replacement clutches but they have been recorded, even with 3 attempts of clutches by a single female.[2][16][114] The females does most but not all of the incubating, doing so for a total of 33–35 days.[2][16] The female remains at the nest brooding the young in the early stages with the male bringing all prey. At about 8–12 days, both the male and female will bring prey but female continues to do all feeding until the young can tear up their own prey.[16]
Development of young[]

Once hatching commences, it may take 48 hours for the chick to chip out. Hatching may take place over 3–7 days, with new hatchlings averaging about 45 g (1.6 oz) in body mass.[2][16][4] Often the youngest nestling dies from starvation, especially in broods of three or more.[16] In nestlings, the first down replaces by longer, coarser down at about 7 days of age with the first proper feathers appearing at 12 to 15 days. The young are nearly fully feathered rather than downy at about a month of age and can start to feed themselves as well. The first attempts to leave the nest are often at about 40–50 days, averaging usually 40–45 in nominate buzzards in Europe, but more quickly on average at 40–42 in vulpinus. Fledging occurs typically at 43–54 days but in extreme cases at as late 62 days. Sexual dimorphism is apparent in European fledglings, as females often scale about 1,000 g (2.2 lb) against 780 g (1.72 lb) in males.[16][4][210] After leaving the nest, buzzards generally stay close by, but with migratory ones there is more definitive movement generally southbound. Full independence is generally sought 6 to 8 weeks after fledging. 1st year birds generally remain in wintering area for following summer but then return to near area of origin but then migrate south again without breeding.[2][16][4] Radio-tracking suggests that most dispersal, even relatively early dispersals, by juvenile buzzards is undertaken independently rather than via exile by parents, as has been recorded in some other birds of prey.[211] In common buzzards, generally speaking, siblings stay quite close to each other after dispersal from their parents and form something of a social group, although parents usually tolerate their presence on their territory until they are laying another clutch. However, the social group of siblings disbands at about a year of age.[210][83] Juvenile buzzards are subordinate to adults during most encounters and tend to avoid direct confrontations and actively defended territories until they are of appropriate age (usually at least 2 years of age).[4] This was the case as well for steppe buzzard juveniles wintering in southern Africa, although in some cases juveniles were able to successfully steal prey from adults there.[212]

Breeding success rates[]
Numerous factors may weigh into the breeding success of common buzzards. Chiefly among these are prey populations, habitat, disturbance and persecution levels and innerspecies competition.[16] In Germany, intra- and interspecific competition, plumage morph, laying date, precipitation levels and anthropogenic disturbances in the breeding territory, in declining order, were deemed to be the most significant bearers of breeding success.[192] In an accompanying study, it was found that a mere 17% of adult birds of both sexes present in a German study area produced 50% of offspring, so breeding success may be lower than perceived and many adult buzzards for unknown causes may not attempt to breed at all.[213] High breeding success was detected in Argyll, Scotland, due likely to hearty prey populations (rabbits) but also probably a lower local rate of persecution than elsewhere in the British isles. Here, the mean number of fledglings were 1.75 against 0.82–1.41 in other parts of Britain.[184] It was found in the English Midlands that breeding success both by measure of clutch size and mean number of fledglings, was relatively high thanks again to high prey populations. Breeding success was lower farther from significant stands of trees in the Midlands and most nesting failures that could be determined occurred in the incubation stage, possibly in correlation with predation of eggs by corvids.[91] More significant than even prey, late winter-early spring was found to be likely the primary driver of breeding success in buzzards from southern Norway. Here, even in peak vole years, nesting success could be considerably hampered by heavy snow at this crucial stage. In Norway, large clutches of 3+ were expected only in years with minimal snow cover, high vole populations and lighter rains in May–June.[208] In the Italian Alps, the mean number of fledglings per pair was 1.07.[124] 33.4% of nesting attempts were failures per a study in southwestern Germany, with an average of 1.06 of all nesting attempts and 1.61 for all successful attempt. In Germany, weather conditions and rodent populations seemed to be the primary drivers of nesting success.[204] In Murcia part of Spain contrasted with Biscay to the north, higher levels of interspecific competition from booted eagles and northern goshawks did not appear to negatively affect breeding success due to more ample prey populations (rabbits again) in Murcia than in Biscay.[86]
In the Westphalia area of Germany, it was found that intermediate colour morphs were more productive than those that were darker or lighter.[213] For reasons that are not entirely clear, apparently fewer parasites were found to afflict broods of intermediate plumaged buzzard less so than dark and light phenotypes, in particular higher melanin levels somehow were found to be more inviting to parasitic organism that effect the health of the buzzard's offspring.[214] The composition of habitat and its relation to human disturbance were important variables for the dark and light phenotypes but were less important to intermediate individuals. Thus selection pressures resulting from different factors did not vary much between sexes but varied between the three phenotypes in the population.[215] Breeding success in areas with wild European rabbits was considerably effected by rabbit myxomatosis and rabbit haemorrhagic disease, both of which have heavily depleted wild rabbit population. Breeding success in formerly rabbit-rich areas were recorded to decrease from as much as 2.6 to as little as 0.9 young per pair.[162][216][217] Age of first breeding in several radio-tagged buzzards showed only a single male breeding as early as his 2nd summer (at about a year of age). Significantly more buzzards were found to start breeding at the 3 summer but breeding attempts can be individually erratic given the availability of habitat, food and mates.[218] The mean life expectancy was estimated at 6.3 years in the late 1950s, but this was at a time of high persecution when humans were causing 50–80% of buzzard deaths.[89] In a more modern context with regionally reduced persecution rates, the lifespan expected can be higher (possibly in excess of 10 years at times) but is still widely variable due to a wide variety of factors.[215]
Status[]

The common buzzard is one of the most numerous birds of prey in its range. Almost certainly, it is the most numerous diurnal bird of prey throughout Europe. Conservative estimates put the total population at no fewer than 700,000 pairs in Europe, which are more than twice the total estimates for the next four birds of prey estimated as most common: the Eurasian sparrowhawk (more than 340,000 pairs), the common kestrel (more than 330,000 pairs) and the northern goshawk (more than 160,000 pairs).[5] Ferguson-Lees et al. roughly estimated that the total population of the common buzzard ranges to nearly 5 million pairs but at time was including the now spilit-off species of eastern and Himalayan buzzards in those numbers.[2] These numbers may be excessive but the total population of common buzzards is certain to total well over seven figures.[1][2] More recently, the IUCN estimated the common buzzard (sans the Himalayan and eastern subspecies) to number somewhere between 2.1 and 3.7 million birds, which would put this buzzard one of the most numerous of all accipitrid family members (estimates for Eurasian sparrowhawks, red-tailed hawks and northern goshawks also may range over 2 million).[1] In 1991, other than their absence in Iceland, after having been extent as breeder by 1910, buzzards recolonized Ireland sometime in the 1950s and has increased by the 1990s to 26 pairs.[2][116] Supplemental feeding has reportedly helped the Irish buzzard population to rebound, especially where rabbits have decreased.[58] Most other countries have at least four figures of breeding pairs. As of the 1990s, other countries such as Great Britain, France, Switzerland, Czech Republic, Poland, Sweden, Belarus and the Ukraine all numbered pairs well into five figures, while Germany had an estimated 140,000 pairs and European Russian may have held 500,000 pairs.[2][5] Between 44,000 and 61,000 pairs nested in Great Britain by 2001 with numbers gradually increasing after past persecution, habitat alteration and prey reductions, making it by far the most abundant diurnal raptor there.[219] In Westphalia, Germany, population of Buzzards was shown to nearly triple over the last few decades. The Westphalian buzzards are possibly benefiting from increasingly warmer mean climate, which in turn is increasing vulnerability of voles. However, the rate of increase was significantly greater in males than in females, in part because of reintroduced Eurasian eagle-owls to the region preying on nests (including the brooding mother), which may in turn put undue pressure on the local buzzard population.[220]
At least 238 common buzzards killed through persecution were recovered in England from 1975 to 1989, largely through poisoning. Persecution did not significantly differ at any time due this span of years nor did the persecution rates decrease, nor did it when compared to rates of last survey of this in 1981.[221] While some persecution persists in England, it is probably slightly less common today.[219] The buzzard was found to be the most vulnerable raptor to power-line collision fatalities in Spain probably as it is one of the most common largish birds, and together with the common raven, it accounted for nearly a third of recorded electrocutions.[222] Given its relative abundance, the common buzzard is held as an ideal bioindicator, as they are effected by a range of pesticide and metal contamination through pollution like other raptors but are largely resilient to these at the population levels. In turn, this allows biologists to study (and harvest if needed) the buzzards intensively and their environments without affecting their overall population. The lack of affect may be due to the buzzard's adaptability as well as its relatively short, terrestrially-based food chain, which exposes them to less risk of contamination and population depletions than raptors that prey more heavily on water-based prey (such as some large eagles) or other birds (such as falcons).[223][224][225][226][227] Common buzzards are seldom vulnerable to egg-shell thinning from DDT as are other raptors but egg-shell thinning has been recorded.[228] Other factors that negatively effect raptors have been studied in common buzzards are helminths, avipoxvirus and assorted other viruses.[229][230][231][232]
Gallery[]

At Hamerton Zoo, England

In flight, Devon, England. There are around 40,000 breeding pairs in the United Kingdom

Egg, Collection Museum Wiesbaden

A young buzzard being ringed

Infertile egg of a Buzzard (Buteo buteo).




References[]
Citations[]
- ^ a b c d e f g h i BirdLife International (2020). "Buteo buteo". IUCN Red List of Threatened Species. 2020: e.T61695117A181770099. doi:10.2305/IUCN.UK.2020-3.RLTS.T61695117A181770099.en. Retrieved 13 November 2021.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi Ferguson-Lees, J., & Christie, D. A. (2001). Raptors of the world. Houghton Mifflin Harcourt.
- ^ a b c Bildstein, K. L., & Zalles, J. I. (2005). Old World versus New World long-distance migration in accipiters, buteos, and falcons. Birds of two worlds: the ecology and evolution of migration. Johns Hopkins University Press, Baltimore, Maryland, USA, 154–167.
- ^ a b c d e f g h i j k Cramp, S., & Brooks, D. J. (1992). Handbook of the birds of Europe, the Middle East and North Africa. The birds of the western Palearctic, vol. 2'’. Oxford University Press, Oxford.
- ^ a b c d BirdLife, F. V. B. (2005). Birds in Europe: population estimates, trends and conservation status. British Birds, 98, 269–271.
- ^ Linnaeus, C. (1758). Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata (in Latin). Volume 1. Holmiae (Stockholm): Laurentii Salvii. p. 90.
F. cera pedibusque luteis, corpore fusco, abdomine paludo maculis fuscis.
|volume=has extra text (help) - ^ Lacépède, Bernard Germain de (1799). "Tableau des sous-classes, divisions, sous-division, ordres et genres des oiseux". Discours d'ouverture et de clôture du cours d'histoire naturelle (in French). Paris: Plassan. p. 4. Page numbering starts at one for each of the three sections.
- ^ Mayr, Ernst; Cottrell, G. William, eds. (1979). Check-list of Birds of the World. Volume 1. 1 (2nd ed.). Cambridge, Massachusetts: Museum of Comparative Zoology. p. 361.
- ^ Jobling, James A (2010). The Helm Dictionary of Scientific Bird Names. London: Christopher Helm. pp. 81. ISBN 978-1-4081-2501-4.
- ^ a b Riesing, M. J., Kruckenhauser, L., Gamauf, A., & Haring, E. (2003). "Molecular phylogeny of the genus Buteo (Aves: Accipitridae) based on mitochondrial marker sequences". Molecular Phylogenetics and Evolution. 27 (2): 328–42. doi:10.1016/S1055-7903(02)00450-5. PMID 12695095.CS1 maint: multiple names: authors list (link)
- ^ a b Clark, W. S. (2007). "Taxonomic status of the Forest Buzzard Buteo oreophilus trizonatus" (PDF). Ostrich. 78: 101–104. doi:10.2989/OSTRICH.2007.78.1.16.60. S2CID 6270968.
- ^ Haring, E., Riesing, M. J., Pinsker, W., & Gamauf, A. (1999). Evolution of a pseudo‐control region in the mitochondrial genome of Palearctic buzzards (genus Buteo) . Journal of Zoological Systematics and Evolutionary Research, 37(4), 185–194.
- ^ Siegfried, W. R. (1969). Affinities of the small African and Palaearctic buteos. Ostrich, 40(S1), 41–46.
- ^ a b Clouet, M., & Wink, M. (2000). The buzzards of Cape Verde Buteo (buteo) bannermani and Socotra Buteo (buteo) spp.: first results of a genetic analysis based on nucleotide sequences of the cytochrome b gene. Alauda, 68(1), 55–58.
- ^ Boev, Z., & Kovachev, D. (1998). Buteo spassovi sp. N.–a late Mionece buzzard (Accipitridae, Aves) from SW Bulgaria. Geologica Balcanica, 29(1–2), 125–129.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah Brown, Leslie and Amadon, Dean (1986) Eagles, Hawks and Falcons of the World. The Wellfleet Press. ISBN 978-1555214722.
- ^ a b c CRC Handbook of Avian Body Masses, 2nd Edition by John B. Dunning Jr. (Editor). CRC Press (2008), ISBN 978-1-4200-6444-5.
- ^ Selas, V. (1997). Nest-site selection by four sympatric forest raptors in southern Norway. Journal of Raptor Research, 31, 16–25.
- ^ Zuberogoitia, I., Martínez, J. A., Zabala, J., Martínez, J. E., Castillo, I., Azkona, A., & Hidalgo, S. (2005). Sexing, ageing and moult of Buzzards Buteo buteo in a southern European area. Ringing & Migration, 22(3), 153–158.
- ^ a b c d e James, A. H. (1984). Geographic variation in the buzzard Buteo buteo (Linnaeus, 1758): mid-Atlantic and west Mediterranean islands (Aves: Accipitridae). Beaufortia, 34(4), 101–116.
- ^ Thibault, Jena-Claude; Patrimonio, Olivier; Torre, José (July 1992). "Does the diurnal raptor community of Corsica (Western Mediterranean) show insular characteristics?". Journal of Biogeography. 19 (4): 363–373. doi:10.2307/2845564. eISSN 1365-2699. ISSN 0305-0270. JSTOR 2845564.
- ^ "Steppe Buzzard – Buteo buteo vulpinus (Gloger, 1833)". Avibase.
- ^ a b c Kruckenhauser, L., Haring, E., Pinsker, W., Riesing, M. J., Winkler, H., Wink, M., & Gamauf, A. (2004). Genetic vs. morphological differentiation of Old World buzzards (genus Buteo, Accipitridae). Zoologica Scripta, 33(3), 197–211.
- ^ a b Voous, K. H., Hens, P. A., & Van Marle, J. G. (1948). The distinguishing characters of the Steppe-Buzzard. HF &G. Witherby.
- ^ Dementiev, G. P., & Gladkov, N. A. (1951). Birds of the Soviet Union, vol. 1. Sovetskaya Nauka, Moscow.
- ^ a b Gorney, E., & Yom‐Tov, Y. (1994). Fat, hydration condition, and moult of Steppe Buzzards Buteo buteo vulpinus on spring migration. Ibis, 136(2), 185–192.
- ^ Yosef, R., Tryjanowski, P., & Bildstein, K. L. (2002). Spring migration of adult and immature buzzards (Buteo buteo) through Elat, Israel: timing and body size. Journal of Raptor Research, 36(2), 115–120.
- ^ Benson, C. W., & Irwin, M. P. S. (1967). A contribution to the ornithology of Zambia (No. 1). published.
- ^ Broekhuysen, G. J., & Siegfried, W. R. (1969). Age and moult in the Steppe Buzzard in southern Africa. Ostrich, 40(S1), 223–237.
- ^ "Common Buzzard (Eastern Steppe) – Buteo buteo menetriesi Bogdanov, 1879". Avibase.
- ^ Mackworth-Praed, C. W., & Grant, C. H. B. (1952). Birds of eastern and north eastern Africa (Vol. 1). Longmans, Green.
- ^ Haring, E., Riesing, M. J., Pinsker, W., & Gamauf, A. (1999). Evolution of a pseudo‐control region in the mitochondrial genome of Palearctic buzzards (genus Buteo). Journal of Zoological Systematics and Evolutionary Research, 37(4), 185–194.
- ^ Voous, K. H., & Bijleveld, M. F. I. J. (1964). A note on Himalayan Buzzards, Buteo buteo (Aves). Beaufortia, 11(134), 37–43.
- ^ Aspinall, S. (2001). The Buteo population of Socotra. Falco, 8.
- ^ a b c d e f Forsman, D. (1999). The raptors of Europe and the Middle East: a handbook of field identification. London: T & AD Poyser.
- ^ Dunning, John B. Jr., ed. (1992). CRC Handbook of Avian Body Masses. CRC Press. ISBN 978-0-8493-4258-5.
- ^ del Hoyo, J.; Elliott, A.; Sargatal, J., eds. (1999). Handbook of the Birds of the World. 2. Barcelona: Lynx Edicions. ISBN 978-84-87334-15-3.
- ^ a b c Clark, W. S. (1999). A field guide to the raptors of Europe, the Middle East, and North Africa. Oxford University Press, USA.
- ^ a b c d e Porter, R. F. (1981). Flight identification of European raptors. A&C Black.
- ^ Dittrich, W. (1985). Gefiedervariationen beim Mäusebussard (Buteo buteo) in Nordbayern. Journal für Ornithologie, 126(1), 93–97.
- ^ Schreiber, A., Stubbe, A., & Stubbe, M. (2001). Common Buzzard (Buteo buteo): A raptor with hyperpolymorphic plumage morphs, but low allozyme heterozygosity. Journal für Ornithologie, 142(1), 34–48.
- ^ Rohwer, S., & Paulson, D. R. (1987). The avoidance-image hypothesis and color polymorphism in Buteo hawks. Ornis Scandinavica, 285–290.
- ^ Duff, D. (2006). Has the juvenile plumage of Honey-buzzard evolved to mimic that of Common Buzzard? British Birds, 99, 118–128.
- ^ Elorriaga, J., & Muñoz, A. R. (2013). Hybridisation between the Common Buzzard Buteo buteo buteo and the North African race of Long-legged Buzzard Buteo rufinus cirtensis in the Strait of Gibraltar: prelude or preclude to colonisation? Ostrich, 84(1), 41–45.
- ^ a b c d e Steyn, P. (1983). Birds of prey of southern Africa: Their identification and life histories. Croom Helm, Beckenham (UK). 1983.
- ^ Unwin, M. (2011). The atlas of birds: diversity, behavior, and conservation. Princeton University Press.
- ^ a b Meheretu Yonas; Leirs, H (2019). Raptor perch sites for biological control of agricultural pest rodents. In: Nyssen J., Jacob, M., Frankl, A. (Eds.). Geo-trekking in Ethiopia's Tropical Mountains – The Dogu'a Tembien District. SpringerNature. ISBN 978-3-030-04954-6.
- ^ Harrison, J. A., & Cherry, M. (1997). The atlas of southern African birds (Vol. 1). Johannesburg: BirdLife South Africa.
- ^ Brown, L., Urban, E. K., Newman, K., Woodcock, M., & Hayman, P. (1982). The birds of Africa (Vol. 1, p. 521). London: Academic Press.
- ^ Viter, S. G. (2019). Forestry Activities and Their Influence on the Raptor's Populations in the Southern Forest-Steppe Zone and Steppe Zone (on the Example of the Kharkiv Region of Ukraine). Raptors Conservation, 38.
- ^ Kamarauskaitė, A., Skuja, S., & Treinys, R. (2019). Nesting habitat overlap between the Common Buzzard Buteo buteo and the Lesser Spotted Eagle Clanga pomarina for conservation planning in Natura 2000 sites. Bird Study, 66(2), 224-233.
- ^ Jȩdrzejewski, W., Jȩdrzejewska, B., & Keller, M. (1988). Nest site selection by the buzzard Buteo buteo L. in the extensive forests of eastern Poland. Biological conservation, 43(2), 145–158.
- ^ Penteriani, V. & Faivre, B. (1997). Habitat selection of Common Buzzard (Buteo buteo) in a mountain area (Abruzzo Apennines, Italy). J. Raptor Res., 31(3), 208–212.
- ^ Butet, A., Michel, N., Rantier, Y., Comor, V., Hubert-Moy, L., Nabucet, J., & Delettre, Y. (2010). Responses of common buzzard (Buteo buteo) and Eurasian kestrel (Falco tinnunculus) to land use changes in agricultural landscapes of Western France. Agriculture, ecosystems & environment, 138(3–4), 152–159.
- ^ Tubbs, C. R., & Tubbs, J. M. (1985). Buzzards Buteo buteo and land use in the New Forest, Hampshire, England. Biological Conservation, 31(1), 41–65.
- ^ Gryz, J., & Krauze-Gryz, D. (2019). The Common Buzzard Buteo buteo Population in a Changing Environment, Central Poland as a Case Study. Diversity, 11(3), 35.
- ^ Palomino, D., & Carrascal, L. M. (2007). Habitat associations of a raptor community in a mosaic landscape of Central Spain under urban development. Landscape and Urban Planning, 83(4), 268–274.
- ^ a b Rooney, E., Reid, N., & Montgomery, W. I. (2015). Supplementary feeding increases Common Buzzard Buteo buteo productivity but only in poor‐quality habitat. Ibis, 157(1), 181–185.
- ^ Stenkat, J., Krautwald-Junghanns, M. E., & Schmidt, V. (2013). Causes of morbidity and mortality in free-living birds in an urban environment in Germany. Ecohealth, 10(4), 352–365.
- ^ Panuccio, M., Foschi, F., Todini, A., Baldi, A., Dominicis, N., De Filippis, P., & Palmeri, A. (2019). Better to stay downtown or in the countryside? Raptors wintering in urban and rural Protected Areas of Rome (Central Italy). Raptor Migration.
- ^ Vysochyn, M. O. (2019). Population dynamics and types of habitats at breeding sites of raptors (Falconiformes) of the Donetsk Ridge along a gradient of anthropogenic disturbance. Regulatory Mechanisms in Biosystems, 10(4), 464-469.
- ^ Kopij, G. (2018). Ecological distribution and population densities of raptors in the inner and outer zone of a Central European city. Biological Bulletin of Bogdan Chmelnitskiy Melitopol State Pedagogical University, 8(1).
- ^ Vergara, P. (2010). Time-of-day bias in diurnal raptor abundance and richness estimated by road surveys. Revista Catalana d’Ornitologia, 26, 22–30.
- ^ Spaar, R., & Bruderer, B. (1997). Optimal flight behavior of soaring migrants: a case study of migrating steppe buzzards, Buteo buteo vulpinus. Behavioral Ecology, 8(3), 288–297.
- ^ a b Spaar, R. (1995). Flight behavior of Steppe Buzzards (Buteo buteo vulpinus) during spring migration in southern Israel: a tracking-radar study. Israel Journal of Zoology, 41(3), 489–500.
- ^ Porter, R. F., & Beaman, M. A. S. (1985). A resume of raptor migration in Europe and the Middle East. Conservation Studies on Raptors. ICBP Technical Publication, 5, 237–242.
- ^ a b Wuczyński, A. (2003). Abundance of Common Buzzard (Buteo buteo) in the Central European wintering ground in relation to the weather conditions and food supply. Buteo, 13, 11–20.
- ^ Walls, S. S., & Kenward, R. E. (1998). Movements of radio‐tagged Buzzards Buteo buteo in early life. Ibis, 140(4), 561–568.
- ^ a b c Schindler, S., Hohmann, U., Probst, R., Nemeschkal, H. L., & Spitzer, G. (2012). Territoriality and habitat use of Common Buzzards (Buteo buteo) during late autumn in northern Germany. Journal of Raptor Research, 46(2), 149–157.
- ^ Schindler, S. (2002). Territoriality and habitat-use of wintering Common Buzzards (Buteo buteo) in Schleswig-Holstein, Germany (Doctoral dissertation).
- ^ Nikolov, S., Spasov, S., & Kambourova, N. (2006). Density, number and habitat use of Common Buzzard (Buteo buteo) wintering in the lowlands of Bulgaria. Buteo, 15, 39–47.
- ^ Baltag, E. S., Pocora, V., Sfica, L., & Bolboaca, L. E. (2013). Common Buzzard (Buteo buteo) population during winter season in North-Eastern Romania: the influences of density, habitat selection, and weather. Ornis Fennica, 90(3), 186.
- ^ Kjellén, N., & Roos, G. (2000). Population trends in Swedish raptors demonstrated by migration counts at Falsterbo, Sweden 1942–97. Bird Study, 47(2), 195–211.
- ^ a b Kjellén, N. (1999). Different migration strategies among Swedish Common Buzzards Buteo buteo revealed by the proportion of white birds. Ornis svecica, 9(1), 11–18.
- ^ Panuccio, M., Duchi, A., Lucia, G., & Agostini, N. (2017). Species-specific behaviour of raptors migrating across the Turkish straits in relation to weather and geography. Ardeola, 64(2), 305–325.
- ^ Porter, R., & Willis, I. (1968). The autumn migration of soaring birds at the Bosphorus. Ibis, 110(4), 520–536.
- ^ Leshem, Y. (1985). Israel: an international axis of raptor migration. ICBP Tech. Publ, 5, 243–250.
- ^ Schmitt, M. B., Baur, S., & Von Maltitz, F. (1980). Observations on the Steppe Buzzard in the Transvaal. Ostrich, 51(3), 151–159.
- ^ a b Herremans, M. (2000). The 'chaotic’flight feather moult of the Steppe Buzzard Buteo buteo vulpinus. Bird Study, 47(3), 332–343.
- ^ Martín, B., Onrubia, A., & Ferrer, M. A. (2014). Effects of climate change on the migratory behavior of the Common Buzzard Buteo buteo. Climate Research, 60(3), 187–197.
- ^ Bensusan, K. J., Garcia, E. F., & Cortes, J. E. (2007). Trends in abundance of migrating raptors at Gibraltar in spring. Ardea, 95(1), 83–91.
- ^ Fryer, G. (1986). Notes on the breeding biology of the buzzard. British Birds, 79(1), 18–28.
- ^ a b Walls, S. S., & Kenward, R. E. (2001). Spatial consequences of relatedness and age in buzzards. Animal Behaviour, 61(6), 1069–1078.
- ^ a b c d e f g Redinov, K.А. (2012). Trophic links of the Buzzard in Ukraine. Russian Birds: 155–168.
- ^ a b c d Selas, V., Tveiten, R., & Aanonsen, O. M. (2007). Diet of common buzzards (Buteo buteo) in southern Norway determined from prey remains and video recordings. Ornis Fennica, 84(3), 97.
- ^ a b c d e f g h i Zuberogoitia, I., Martínez, J. E., Martínez, J. A., Zabala, J., Calvo, J. F., Castillo, I., Azkona, A., Iraeta, A. & Hidalgo, S. (2006). Influence of management practices on nest site habitat selection, breeding and diet of the common buzzard Buteo buteo in two different areas of Spain. Ardeola, 53(1), 83–98.
- ^ a b c d Sidorovich, A. A., Ivanovskij, V. V., Sidorovich, V. E., & Solovej, I. A. (2016). Landscape-related variation in the diet composition of the common buzzard (Buteo buteo) in Belarus. Slovak Raptor Journal, 10(1), 65–74.
- ^ Spidsø, T. K., & Selås, V. (1988). Prey selection and breeding success in the common buzzard Buteo buteo in relation to small rodent cycles in southern Norway. Fauna Norvegica, Series C, 11(2), 61–66.
- ^ a b Mebs, T. (1964). Zur Biologie und Populationsdynamik des Mäusebussards (Buteo buteo)(Unter besonderer Berücksichtigung der Abhängigkeit vom Massenwechsel der Feldmaus Microtus arvalis). Journal für Ornithologie, 105(3), 247–306.
- ^ a b Kenward, R. E., Hall, D. G., Walls, S. S., & Hodder, K. H. (2001). Factors affecting predation by buzzards Buteo buteo on released pheasants Phasianus colchicus. Journal of Applied Ecology, 38(4), 813–822.
- ^ a b c d Sim, I. M. W., Cross, A. V., Lamacraft, D. L., & Pain, D. J. (2001). Correlates of Common Buzzard Buteo buteo density and breeding success in the West Midlands. Bird Study, 48(3), 317–329.
- ^ a b Halle, S. (1988). Avian predation upon a mixed community of common voles (Microtus arvalis) and wood mice (Apodemus sylvaticus). Oecologia, 75(3), 451–455.
- ^ a b c d e f g h i j Selås, V. (2001). Predation on reptiles and birds by the common buzzard, Buteo buteo, in relation to changes in its main prey, voles. Canadian Journal of Zoology, 79(11), 2086–2093.
- ^ Andersson, M., & Erlinge, S. (1977). Influence of predation on rodent populations. Oikos, 591–597.
- ^ a b c d e f Graham, I. M., Redpath, S. M., & Thirgood, S. J. (1995). The diet and breeding density of Common Buzzards Buteo buteo in relation to indices of prey abundance. Bird Study, 42(2), 165–173.
- ^ Francksen, R. M.; Whittingham, M. J.; Ludwig, S. C.; Roos, S.; Baines, D. (2017). "Numerical and functional responses of Common Buzzards Buteo buteo to prey abundance on a Scottish grouse moor". Ibis. 159 (3): 541–553. doi:10.1111/ibi.12471.
- ^ a b c d e f g h i Bijlsma, R. G. (2012). Reptielen als voer voor jonge Buizerds Buteo buteo. De takkeling, 20(2), 133–144.
- ^ a b c d e f g h Šotnár, K., & Obuch, J. (2009). Feeding ecology of a nesting population of the Common Buzzard (Buteo buteo) in the Upper Nitra Region, Central Slovakia. Slovak Raptor Journal, 3, 13–20.
- ^ a b c d e f g h i Chernichko, I.I. & Davygora, A.V. (editors). (2012). Buzzards of North Eurasia: Distribution, Population Status, Biology. Proceedings of the 6th International Conference on Birds of Prey and Owls of North Eurasia, Kryvyi Rih, 27–30 September 2012. – Kryvyi Rih: Press- 272 p.
- ^ a b Costa, A. S. C. (2017). Estudo da dieta de Águia-d'asa-redonda (Buteo buteo [Linnaeus, 1758]) e de Águia-calçada (Hieraaetus pennatus [Gmelin, 1788]) em montados de sobro do Alto-Alentejo (Doctoral dissertation).
- ^ McComb, A. M. (2004). Raptor Predation Attempts on Grey Squirrels Sciurus carolinensis (Gmelin). The Irish Naturalists' Journal, 27(12), 483–484.
- ^ Lima, S. L., Valone, T. J., & Caraco, T. (1985). Foraging-efficiency-predation-risk trade-off in the grey squirrel. Animal Behaviour, 33(1), 155–165.
- ^ Ramos-Lara, N., Koprowski, J. L., Kryštufek, B., & Hoffmann, I. E. (2014). Spermophilus citellus (Rodentia: sciuridae). Mammalian Species, 46(913), 71–87.
- ^ Matrosova, V. A., Schneiderová, I., Volodin, I. A., & Volodina, E. V. (2012). Species-specific and shared features in vocal repertoires of three Eurasian ground squirrels (genus Spermophilus). Acta Theriologica, 57(1), 65–78.
- ^ Makomaska-Juchiewicz, M., & Baran, P. (2012). Monitoring gatunków zwierząt. Przewodnik metodyczny. Część III. GIOŚ, Warszawa, 292–496.
- ^ a b c Palomar, C. J. P. (2005). El ratonero común (Buteo buteo insularum) en Fuerteventura, islas Canarias (Aves, Accipitridae). Vieraea, 33, 1–7.
- ^ a b c Steiner, H. (1999). Der Mäusebussard (Buteo buteo) als Indikator für Struktur und Bodennutzung des ländlichen Raumes: produktivität im heterogenen Habitat, Einfluß von Nahrung und Witterung und Vergleiche zum Habicht (Accipiter gentilis) (Vol. 62). Biologiezentrum des OÖ. Landesmuseums.
- ^ Åberg, B., & Hungate, F. P. (Eds.). (2013). Radioecological Concentration Processes: Proceedings of an International Symposium Held in Stockholm, 25–29 April 1966. Elsevier.
- ^ Chapuis, J.L. (2006). Tamias sibiricus. Delivering Alien Invasive Species Inventories.
- ^ a b c Pinowski, J., & Ryszkowski, L. (1962). The Buzzard’s versatility as a predator. British Birds, 55, 470–475.
- ^ Toyran, K., & Albayrak, İ. (2009). Contribution to the Biological Characteristics of Allactaga williamsi Thomas, 1897 in Kırıkkale Province (Mammalia: Rodentia) . Uluslararası Doğa ve Mühendislik Bilimleri Dergisi, (1), 13–17.
- ^ Nachtigall, W. (1996). Siebenschlafer (Glis glis; Mammalia: Rodentia: Gliridae) als Beute des Mausebussards (Buteo buteo, Aves: Falconiformes: Accipitridae). (Beitrage zur Saugetierfauna Sachsens, No. 1). Faunistische Abhandlungen, 20: 320.
- ^ Gwynn, L. (2015). The identity, origin and impact of a 'new' buzzard species breeding in South Africa (Doctoral dissertation, University of Cape Town).
- ^ a b c d e f g h i j Swann, R. L., & Etheridge, B. (1995). A comparison of breeding success and prey of the Common Buzzard Buteo buteo in two areas of northern Scotland. Bird Study, 42(1), 37–43.
- ^ a b Swan, G. (2011). Spatial variation in the breeding success of the common buzzard Buteo buteo in relation to habitat type and diet (Doctoral dissertation, Department of Life Sciences, Silwood Park, Imperial College London).
- ^ a b c d Rooney, E., & Montgomery, W. I. (2013). Diet diversity of the Common Buzzard (Buteo buteo) in a vole-less environment. Bird Study, 60(2), 147–155.
- ^ a b c d e Manosa, S. & Cordero, P.J. (1992). SEASONAL AND SEXUAL VARIATION IN THE DIET OF THE COMMON BUZZARD IN NORTHEASTERN SPAIN. J. Raptor Res., 26(4), 235–238.
- ^ Reif, V., Tornberg, R., Jungell, S., & Korpimäki, E. (2001). Diet variation of common buzzards in Finland supports the alternative prey hypothesis. Ecography, 24(3), 267–274.
- ^ Francksen, R. M., Whittingham, M. J., & Baines, D. (2016). Assessing prey provisioned to Common Buzzard Buteo buteo chicks: a comparison of methods. Bird Study, 63(3), 303–310.
- ^ Skierczyński, M. (2006). Food niche overlap of three sympatric raptors breeding in agricultural landscape in Western Pomerania region of Poland.
- ^ a b Scillitani, G. (1999). Dieta della poiana, Buteo buteo (L., 1758) in provincia di Bari. Avocetta, 23: 169.
- ^ Korpimäki, E., & Norrdahl, K. (1989). Avian predation on mustelids in Europe 1: occurrence and effects on body size variation and life traits. Oikos, 205–215.
- ^ Selva, N., Jedrzejewska, B., Jedrzejewski, W., & Wajrak, A. (2003). Scavenging on European bison carcasses in Bialowieza primeval forest (eastern Poland). Ecoscience, 10(3), 303–311.
- ^ a b c d e f Sergio, F., Boto, A., Scandolara, C., & Bogliani, G. (2002). Density, nest sites, diet, and productivity of Common Buzzards (Buteo buteo) in the Italian pre-Alps. Journal of Raptor Research, 36(1), 24–32.
- ^ Parrott, D. (2015). Impacts and management of common buzzards Buteo buteo at pheasant Phasianus colchicus release pens in the UK: a review. European journal of wildlife research, 61(2), 181–197.
- ^ Boonman, M. & de Vrieslaan, H. (1992). [Buzzard Buteo buteo examines a nest of a Grey Heron Ardea cinerea]. Limosa, 65: 29–30.
- ^ Johnsgard, P. A. (1983). Cranes of the World: Black-necked Crane (Grus nigricollis). Cranes of the World, by Paul Johnsgard, Vol. 5.
- ^ Monteiro, L. R., & Furness, R. W. (1998). Speciation through temporal segregation of Madeiran storm petrel (Oceanodroma castro) populations in the Azores? Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 353(1371), 945–953.
- ^ Ramos, J. A., Moniz, Z., Solá, E., & Monteiro, L. R. (2003). Reproductive measures and chick provisioning of Cory's Shearwater Calonectris diomedea borealis in the Azores. Bird Study, 50(1), 47–54.
- ^ Hoi, H., Hoi, C., Kristofik, J., & Darolova, A. (2002). Reproductive success decreases with colony size in the European bee-eater. Ethology Ecology & Evolution, 14(2), 99–110.
- ^ Martin, J. (1990). Amphibians and reptiles as prey of birds in southwestern Europe. Smithsonian Herpetological Information Service.
- ^ Bustamante, J. (1985). Alimentación del ratonero común (Buteo buteo, L. 1758) en el Norte de Espana. Donana, Acta Vertebrata, 12, 51–62.
- ^ Meek, R. (2013). Post hibernation movements in an aspic viper, Vipera aspis. Herpetological Bulletin, 125, 22–24.
- ^ Ettling, J. A., Aghasyan, L. A., Aghasyan, A. L., & Parker, P. G. (2013). Spatial ecology of Armenian Vipers, Montivipera raddei, in a human-modified landscape. Copeia, 2013(1), 64–71.
- ^ Spellerberg, I. F. (1975). The grass snake in Britain. Oryx, 13(2), 179–184.
- ^ Guthrie, J. E. (1932). Snakes versus birds; birds versus snakes. The Wilson Bulletin, 44(2), 88–113.
- ^ Ash, C. P., & Atkins, J. D. (2009). Birds of Ethiopia and Eritrea: an atlas of distribution. A&C Black.
- ^ Irwin, M. P. S. (1981). The birds of Zimbabwe. Quest Pub.
- ^ Long, R. C. (1960). The birds of the Port Herald District.(Part I). Ostrich, 31(3), 85–104.
- ^ Neuvel, J. & Winters, B. (1987). [Buzzard Buteo buteo eats eel]. Limosa, 60: 149.
- ^ Maczkowski, K. & Tryjanowski, P. (1993). Carp (Cyprinus carpio) in the food of buzzard (Buteo buteo). Przeglad Przyrodniczy, 42: 76.
- ^ Davis, T. A. W. (1960). Kestrel pellets at a winter roost. Brit. Birds, 53(7), 281–284.
- ^ Redpath, S. M., Clarke, R., Madders, M., & Thirgood, S. J. (2015). Assessing raptor diet: comparing pellets, prey remains, and observational data at hen harrier nests.
- ^ Lohmus, A., & Väli, Ü. (2004). The effects of habitat quality and female size on the productivity of the lesser spotted eagle Aquila pomarina in the light of the alternative prey hypothesis. Journal of Avian Biology, 35(5), 455–464.
- ^ Meunier, F. D., Verheyden, C., & Jouventin, P. (2000). Use of roadsides by diurnal raptors in agricultural landscapes. Biological Conservation, 92(3), 291–298.
- ^ Bergman, G. (1961). The food of birds of prey and owls in Fenno-Scandia. British Birds, 54, 307–320.
- ^ Jankowiak, L., & Tryjanowski, P. (2013). Cooccurrence and food niche overlap of two common predators (red fox Vulpes vulpes and common buzzard Buteo buteo) in an agricultural landscape. Turkish Journal of Zoology, 37(2), 157–162.
- ^ Sylven, M. (1978). Interspecific relations between sympatrically wintering Common Buzzards Buteo buteo and Rough-legged Buzzards Buteo lagopus. Ornis Scandinavica, 197–206.
- ^ Kassinis, N. (2009). Long-legged buzzard Buteo rufinus rufinus breeding distribution and abundance in Cyprus. Avocetta, 33, 75–78.
- ^ Bakaloudis, D. E., Iezekiel, S., Vlachos, C. G., Bontzorlos, V. A., Papakosta, M., & Birrer, S. (2012). Assessing bias in diet methods for the Long-legged Buzzard Buteo rufinus. Journal of Arid Environments, 77, 59–65.
- ^ Krüger, O. (2002). Analysis of nest occupancy and nest reproduction in two sympatric raptors: common buzzard Buteo buteo and goshawk Accipiter gentilis. Ecography, 25(5), 523–532.
- ^ Krüger, O. (2002). Interactions between common buzzard Buteo buteo and goshawk Accipiter gentilis: trade‐offs revealed by a field experiment. Oikos, 96(3), 441–452.
- ^ Taylor, K., Hudson, R., & Horne, G. (1988). Buzzard breeding distribution and abundance in Britain and Northern Ireland in 1983. Bird Study, 35(2), 109–118.
- ^ Kostrzewa, A. (1991). Interspecific interference competition in three European raptor species. Ethology Ecology & Evolution, 3(2), 127–143.
- ^ a b Hakkarainen, H., Mykrä, S., Kurki, S., Tornberg, R., & Jungell, S. (2004). Competitive interactions among raptors in boreal forests. Oecologia, 141(3), 420–424.
- ^ Goszczyński, J. (1997). Density and productivity of Common Buzzard Buteo buteo and goshawk Accipiter gentilis populations in Rogów, Central Poland. Acta Ornithologica, 32(2), 149–155.
- ^ Gatto, Angela E.; Grubb, Teryl G.; Chambers, Carol L. (2006). "Red-tailed Hawk dietary overlap with Northern Goshawks on the Kaibab Plateau, Arizona". Journal of Raptor Research. 39 (4): 439–444.
- ^ Boal, C. W. (2005). "Productivity and mortality of northern goshawks in Minnesota". J. Raptor Res. 39 (3): 222–228.
- ^ Luttich, S., Rusch, D. H., Meslow, E. C., & Keith, L. B. (1970). "Ecology of Red-Tailed Hawk Predation in Alberta". Ecology. 51 (2): 190–203. doi:10.2307/1933655. JSTOR 1933655.CS1 maint: multiple names: authors list (link)
- ^ Sergio, F., & Boto, A. (1999). Nest dispersion, diet, and breeding success of Black Kites (Milvus migrans) in the Italian pre-Alps. Journal of Raptor Research, 33, 207–217.
- ^ Meyburg, B. U. (1973). Studies of less familiar birds: 172. Lesser Spotted Eagle. British Birds.
- ^ a b Moore, N. W. (1957). The past and present status of the Buzzard in the British Isles. British Birds, 50, 173–197.
- ^ Dixon, N., & Gibbs, A. (2018). Extreme territorial aggression by urban Peregrine Falcons toward Common Buzzards in South-West England. Ornis Hungarica, 26(2), 232-242.
- ^ O'Connor, T. P. (1993). Birds and the scavenger niche. Archaeofauna, (2).
- ^ a b Sergio, F., & Hiraldo, F. (2008). Intraguild predation in raptor assemblages: a review birds. Ibis, 150, 132–145.
- ^ a b Björklund, H., Santangeli, A., Blanchet, F. G., Huitu, O., Lehtoranta, H., Lindén, H., Valkama, J. & Laaksonen, T. (2016). Intraguild predation and competition impacts on a subordinate predator birds. Oecologia, 181(1), 257–269.
- ^ Weiss, N., & Yosef, R. (2010). Steppe Eagle (Aquila nipalensis) Hunts a Eurasian Buzzard (Buteo buteo vulpinus) While in Migration over Eilat, Israel. Journal of Raptor Research, 44(1), 77–79.
- ^ Todd, D.M. (1980). Golden Eagle killing buzzard. British Birds, 73: 536–537.
- ^ Chavko, J., Danko, Š., Obuch, J., & Mihók, J. (2007). The food of the Imperial Eagle (Aquila heliaca) in Slovakia. Slovak Raptor Journal, 1, 1–18.
- ^ Horváth, M., Solti, B., Fatér, I., Juhász, T., Haraszthy, L., Szitta, T., Bállok, Z. & Pásztory-Kovács, S. (2018). Temporal changes in the diet composition of the Eastern Imperial Eagle (Aquila heliaca) in Hungary. Ornis Hungarica, 26(1), 1–26.
- ^ Gradoz, P. (1996). [Common Buzzard Buteo buteo captured in flight by a White-tailed Eagle Haliaetus albicilla.] 62: 165–166.
- ^ Resano, J., Bayle, P., Real, J., Hernández, A., Vincent-Martin, N. & Ravayrol, A. (2012). Analyse du régime alimentaire de l’Aigle de Bonelli Hieraaetus fasciatus (Vieillot, 1822) pendant la saison de reproduction 2010 en France. Université de Barcelone – Equip de Biologia de la Conservació, 1: 95–101.
- ^ Dementavičius, D. (2004). Common Buzzard (Buteo buteo) and White-tailed Eagle (Haliaeetus albicilla): breeding parasitism or atypical feeding behaviour? Acta Zoologica Lituanica, 14(1), 76–79.
- ^ Literak, I., & Mraz, J. (2011). Adoptions of young common buzzards in white-tailed sea eagle nests. The Wilson Journal of Ornithology, 123(1), 174–176.
- ^ Kamarauskaitė, A., Dementavičius, D., Skuja, S., Dagys, M., & Treinys, R. (2020). Interaction between the White-tailed Eagle and Common Buzzard estimated by diet analysis and brood defence behaviour. Ornis Fennica, 97(1), 26-37.
- ^ Dementavičius, D., Rumbutis, S., Virbickas, T., Vaitkuvienė, D., Dagys, M., & Treinys, R. (2020). Spatial and temporal variations in the White-tailed Eagle Haliaeetus albicilla breeding diet revealed by prey remains. Bird Study, 67(2), 206-216.
- ^ Lourenço, R., Rabaça, J. E., Delgado, M. D. M., & Penteriani, V. (2009). Intraguild predation by a European top predator–the Eagle Owl (Bubo bubo).
- ^ Chakarov, N., & Krüger, O. (2010). Mesopredator release by an emergent superpredator: a natural experiment of predation in a three level guild. PLOS ONE, 5(12), e15229.
- ^ Zemanová, K. (2009). Potrava výra velkého na střední Moravě.
- ^ Bayle, P., & Schauls, R. (2011). Biologie de quatre couples de grand-duc d’Europe Bubo bubo au Luxembourg. Bull. Soc. Nat. luxemb, 112, 51.
- ^ Schindler, S. (2002). Territoriality and habitat-use of wintering Common Buzzards (Buteo buteo) in Schleswig-Holstein, Germany (Doctoral dissertation, uniwien).
- ^ Avery, G., Robertson, A. S., Palmer, N. G., & Prins, A. J. (1985). Prey of giant eagle owls in the de Hoop nature reserve, Cape province, and some observations on hunting strategy. Ostrich, 56(1–3), 117–122.
- ^ Byshnev, I.I. (2002). Interesting case of aggressive interaction between Common Buzzard (Buteo buteo) and Ural Owl (Strix uralensis). Subbuteo, 5: 46.
- ^ a b c Austin, G. E., & Houston, D. C. (1997). The breeding performance of the Buzzard Buteo buteo in Argyll, Scotland and a comparison with other areas in Britain. Bird Study, 44(2), 146–154.
- ^ Tubbs, C. R. (1972). Analysis of nest record cards for the Buzzard. Bird Study, 19(2), 97–104.
- ^ Sarà, M., Campobello, D., & Zanca, L. (2012). Effects of nest and colony features on lesser kestrel (Falco naumanni) reproductive success. Avian Biology Research, 5(4), 209–217.
- ^ Mikkola, H. (1976). Owls killing and killed by other owls and raptors in Europe. British Birds, 69, 144–154.
- ^ Van Nieuwenhuyse, D., Genot, J. C., & Johnson, D. H. (Eds.). (2008). The little owl: conservation, ecology and behavior of Athene noctua. Cambridge University Press, The Edinburgh Building, Shaftesbury Road Cambridge CB 2 2 RU UK.
- ^ Barnard. C.C. (1981). Buzzard preying on Short-eared Owl. British Birds, 74: 226.
- ^ Voous, K.H. 1988. Owls of the Northern Hemisphere. The MIT Press, 0262220350.
- ^ Sunde, P. (2005). Predators control post‐fledging mortality in tawny owls, Strix aluco. Oikos, 110(3), 461–472.
- ^ a b Krüger, O. (2004). The importance of competition, food, habitat, weather and phenotype for the reproduction of Buzzard Buteo buteo. Bird Study, 51(2), 125–132.
- ^ Novikova, L. (2009). Number and Distribution of the Common Buzzard in the Kerzhenskiy State Nature Reserve, Russia. Raptors Conservation, (16).
- ^ Dare, P. J., & Barry, J. T. (1990). Population size, density and regularity in nest spacing of Buzzards Buteo buteo in two upland regions of North Wales. Bird Study, 37(1), 23–29.
- ^ a b Rodríguez, B., Siverio, F., Rodríguez, A., Siverio, M., Hernández, J. J., & Figuerola, J. (2010). Density, habitat selection and breeding biology of Common Buzzards Buteo buteo in an insular environment. Bird Study, 57(1), 75–83.
- ^ Tzortzakaki, O., Simaiakis, S., & Xirouchakis, S. (2012). Abundance of common buzzards (Buteo buteo) in olive monocultures in the island of Crete. Journal of Biological Research, 17, 44.
- ^ Cerasoli, M., & Penteriani, V. (1996). Common Buzzards (Buteo buteo) in Central Italy. J. Raptor Res., 30(3), 130–135.
- ^ Šotnár, K., & Topercer, J. (2009). Estimating density, population size and dynamics of Common Buzzard (Buteo buteo) in the West Carpathian region by a new method. Slovak Raptor Journal, 3, 1–12.
- ^ Voříšek, P. (2000). An extremely high population density of common buzzard (Buteo buteo) in Biosphere Reserve Pálava (Czech Republic) and its possible causes. Buteo, 11, 51–56.
- ^ Forsman, D., & Solonen, T. (1984, January). Censusing breeding raptors in southern Finland: methods and results. In Annales Zoologici Fennici (pp. 317–320). Finnish Academy of Sciences, Societas Scientiarum Fennica, Societas pro Fauna et Flora Fennica and Societas Biologica Fennica Vanamo.
- ^ Negro, J. J., & Galván, I. (2018). Behavioural Ecology of Raptors. In Birds of Prey (pp. 33–62). Springer, Cham.
- ^ Simmons, R. E., & Mendelsohn, J. M. (1993). A critical review of cartwheeling flights of raptors. Ostrich, 64(1), 13–24.
- ^ Barrientos, R., & López-Darias, M. (2006). A case of a polyandrous trio of Eurasian buzzards (Buteo buteo) on Fuerteventura Island, Canary Islands. Journal of Raptor Research, 40(4), 305–306.
- ^ a b Rockenbauch, D. (1975). Zwölfjährige Untersuchungen zur Ökologie des Mäusebussards (Buteo buteo) auf der Schwäbischen Alb. Journal für Ornithologie, 116(1), 39–54.
- ^ Austin, G. E. (1992). The distribution and breeding performance of the buzzard Buteo buteo in relation to habitat: an application using remote sensing and geographical information systems (Doctoral dissertation, University of Glasgow).
- ^ Jonkers, D. A., & Roodhart, J. (2003). Grondnest van Buizerd Buteo buteo op bruggetje. De takkeling, 11(3), 234–236.
- ^ Castillo-Gómez, C., & Moreno-Rueda, G. (2011). A record of a Common Buzzard (Buteo buteo) nesting in an abandoned building. Journal of Raptor Research, 45(3), 275–277.
- ^ a b Selas, V. (2001). Breeding density and brood size of Common Buzzard Buteo buteo in relation to snow cover in spring. Ardea, 89(3), 471–479.
- ^ Holdsworth, M. (1971). Breeding biology of Buzzards at Sedbergh during 1937–67. British Birds, 64, 412–420.
- ^ a b Tyack, A. J., Walls, S. S., & Kenward, R. E. (1998). Behaviour in the post‐nestling dependence period of radio‐tagged Common Buzzards Buteo buteo. Ibis, 140(1), 58–63.
- ^ Walls, S. S., Mañosa, S., Fuller, R. M., Hodder, K. H., & Kenward, R. E. (1999). Is early dispersal enterprise or exile? Evidence from radio-tagged buzzards. Journal of Avian Biology, 407–415.
- ^ Herremans, M. (2001). Age-related differences in abundance and habitat occupation of Steppe Buzzards, Buteo buteo vulpinus, in the Western Cape Province, South Africa. Ostrich-Journal of African Ornithology, 72(1–2), 105–108.
- ^ a b Krüger, O., & Lindström, J. (2001). Lifetime reproductive success in common buzzard, Buteo buteo: from individual variation to population demography. Oikos, 93(2), 260–273.
- ^ Chakarov, N., Boerner, M., & Krüger, O. (2008). Fitness in common buzzards at the cross‐point of opposite melanin–parasite interactions. Functional Ecology, 22(6), 1062–1069.
- ^ a b Krüger, O. (2002). Dissecting common buzzard lifespan and lifetime reproductive success: the relative importance of food, competition, weather, habitat and individual attributes. Oecologia, 133(4), 474–482.
- ^ Moore, N. W. (1956). Rabbits, buzzards and hares. Two studies on the indirect effects of myxomatosis. Les conséquences biologiques dues à la présence de la myxomatose.
- ^ Lees, A. C., & Bell, D. J. (2008). A conservation paradox for the 21st century: the European wild rabbit Oryctolagus cuniculus, an invasive alien and an endangered native species. Mammal Review, 38(4), 304–320.
- ^ Davis, P. E., & Davis, J. E. (1992). Dispersal and age of first breeding of buzzards in central Wales. British Birds, 85(11), 578–587.
- ^ a b Clements, R. (2002). The Common Buzzard in Britain. British Birds, 95, 377–383.
- ^ Jonker, R. M., Chakarov, N., & Krüger, O. (2014). Climate change and habitat heterogeneity drive a population increase in Common Buzzards Buteo buteo through effects on survival. Ibis, 156(1), 97–106.
- ^ Elliott, G. D., & Avery, M. I. (1991). A review of reports of buzzard persecution 1975–1989. Bird Study, 38(1), 52–56.
- ^ Janss, G. F. (2000). Avian mortality from power lines: a morphologic approach of a species-specific mortality. Biological Conservation, 95(3), 353–359.
- ^ Naccari, C., Cristani, M., Cimino, F., Arcoraci, T., & Trombetta, D. (2009). Common buzzards (Buteo buteo) bio-indicators of heavy metals pollution in Sicily (Italy). Environment international, 35(3), 594–598.
- ^ Carneiro, M., Colaço, B., Brandão, R., Ferreira, C., Santos, N., Soeiro, V., & Lavín, S. (2014). Biomonitoring of heavy metals (Cd, Hg, and Pb) and metalloid (As) with the Portuguese common buzzard (Buteo buteo). Environmental monitoring and assessment, 186(11), 7011–7021.
- ^ Jager, L. P., Rijnierse, F. V., Esselink, H., & Baars, A. J. (1996). Biomonitoring with the Buzzard Buteo buteo in the Netherlands: heavy metals and sources of variation. Journal für Ornithologie, 137(3), 295–318.
- ^ Manosa, S., Mateo, R., Freixa, C., & Guitart, R. (2003). Persistent organochlorine contaminants in eggs of northern goshawk and Eurasian buzzard from northeastern Spain: temporal trends related to changes in the diet. Environmental Pollution, 122(3), 351–359.
- ^ Tubbs, C. R. (1967). Population study of Buzzards in the New Forest during 1962–66. British Birds, 60(10), 381–395.
- ^ Ratcliffe, D. A. (1967). Decrease in eggshell weight in certain birds of prey. Nature, 215(5097), 208.
- ^ Santoro, M., Tripepi, M., Kinsella, J. M., Panebianco, A., & Mattiucci, S. (2010). Helminth infestation in birds of prey (Accipitriformes and Falconiformes) in Southern Italy. The Veterinary Journal, 186(1), 119–122.
- ^ Shirazi, S., Hesaraki, S., Mostafaei, T. S., & Davoodi, J. (2014). First report on Centrorhynchus aluconis in common buzzard (Buteo buteo) in northwest Iran. Acta Veterinaria, 64(2), 276–280.
- ^ ÖZMEN, Ö., & DORRESTEIN, G. M. (2002). Avipox in a common buzzard (Buteo buteo). Turkish Journal of Veterinary and Animal Sciences, 26(5), 1193–1195.
- ^ Frölich, K., Prusas, C., Schettler, E., & Hafez, H. M. (2002). Antibodies to adenoviruses in free-living common buzzards from Germany. Journal of wildlife diseases, 38(3), 633–636.
General sources[]
- Mullarney, Killian; Svensson, Lars; Zetterstrom, Dan; Grant, Peter (1999). Collins Bird Guide. London: HarperCollins. ISBN 978-0-00-219728-1.
- Sinclair, Ian; Hockey, Phil; Tarboton, Warwick (2002). SASOL Birds of Southern Africa. Struik. ISBN 978-1-86872-721-6.
External links[]
| Wikimedia Commons has media related to Buteo buteo. |
| Wikispecies has information related to Buteo buteo. |
- Steppe Buzzard species text in The Atlas of Southern African Birds
- Madeira Birds: Buzzard. Page about the controversial subspecies harterti. Retrieved 28 November 2006.
- Ageing and sexing (PDF; 4.2 MB) by Javier Blasco-Zumeta & Gerd-Michael Heinze
- Feathers of Common Buzzard (Buteo buteo)
- BirdLife species factsheet for Buteo buteo
- "Buteo buteo". Avibase.

- "Eurasian buzzard media". Internet Bird Collection.
- Common buzzard photo gallery at VIREO (Drexel University)
- Audio recordings of Common buzzard on Xeno-canto.
| Authority control |
|---|
- IUCN Red List least concern species
- Buteo
- Birds of Africa
- Birds of Asia
- Birds of Europe
- Birds of Macaronesia
- Birds described in 1758
- Taxa named by Carl Linnaeus