Conocephalum
| Conocephalum | |
|---|---|

| |
| Conocephalum conicum | |
| Scientific classification | |
| Kingdom: | Plantae |
| Division: | Marchantiophyta |
| Class: | Marchantiopsida |
| Order: | Marchantiales |
| Family: | Conocephalaceae Müll.Frib. ex Grolle |
| Genus: | Conocephalum Hill, 1773, corr. Wiggers 1780, >nom. et orth. cons. |
Conocephalum is a genus of complex thalloid liverworts in the order Marchantiales and is the only extant genus in the family Conocephalaceae.[1][2] Some species of Conocephalum are assigned to the Conocephalum conicum complex, which includes several cryptic species.[1] Cryptic speciation refers to species which display clear genetic differences but show little or no morphological differences.[3] Species of Conocephalum are relatively common[4] and widely distributed throughout North America, Europe and East Asia.[5] Conocephalum species are large liverworts with distinct patterns on the upper thallus, giving the appearance of snakeskin.[6] The name Conocephalum conicum refers to the cone-shaped archegoniophores, which bear sporangia.[7] Common names include snakeskin liverwort, great scented liverwort[4][8] and cat-tongue liverwort.[9]
Classification and Taxonomy[]
Some species of Conocephalum are placed in the Conocephalum conicum complex, which includes several cryptic species.[1] Consequently, it has been challenging to identify the exact number of species in this genus.[10] Cryptic species refers to a species which demonstrates a genetic difference but lacks morphological differences. Within liverworts, cryptic species are suggested to be related to both geographical disjunction and to reproductive biology in combination with isolation and habitat differentiation.[3]
Molecular research has indicated that Conocephalum comprises a complex of six cryptic species (A, C, F, J, L and S).[3][11] In 2005 C. conicum cryptic species S was described as a separate species, Conocephalum salebrosum.[1][3][12] C. salebrosum has a wider distribution and is present in North America, in contrast to C. conicum.[1][6][12] More recent examinations of the Conocephalum conicum complex in Japan and Taiwan have identified three new species within Conocephalum, C. orientalis, C. purpureorubum and C. toyotae, which were formerly described as C. conicum J, F, and R.[2]
Species[]
- Conocephalum conicum complex – includes several cryptic species:[1][2]
- Conocephalum conicum
- Conocephalum salebrosum
- Conocephalum orientalis
- Conocephalum purpureorubum
- Conocephalum toyotae
- Conocephalum supradecompositum
Distribution[]
Species of Conocephalum are distributed throughout North America, Europe and East Asia.[5][10][13] C. salebrosum displays the widest distribution and is found throughout North America,[4][10][12] Europe and Asia.[14] In North America, C. salebrosum occurs throughout Canada and parts of the United States. C. salebrosum has also been recorded in Russia. [12] C. conicum is found throughout Europe[10][13] and has been recorded in Norway, Finland, Great Britain, Ireland, Belgium, France, Germany, Czech Republic, Austria, Hungary, Romania, Italy, Portugal, Spain, Croatia, Bulgaria, Greece, Ukraine, Poland and Russia.[15] Conocephalum supradecompositum is more restricted in its distribution and is mainly found in China and Japan.[16][17] Regarding the most recently described species, C. purpureorubum has been observed in Japan, China, Taiwan and South Korea and C. orientalis has been found in Japan and Taiwan.[2][13]
Habitat[]
Species of Conocephalum often occur in moist and shaded habitats.[1] Conocephalum species also often grow in specialized micro-habitats near both running or standing water.[14] C. conicum is often found in open woodlands, sandy banks, wet rocks and cliffs and moist soils.[7] C. conicum is also strongly associated with calcareous substrates.[12] C. salebrosum occurs in moist, shaded and calcareous habitats.[4][12][14][18][19] In contrast to C. conicum, C. salebrosum is more tolerant of desiccation.[12][13][14][18][19]
Morphology[]


C. conicum and C. salebrosum share some similarities in morphological characteristics, in addition to having their own characteristics which help distinguish the two species.[19]

Gametophyte[]
General morphology[]
The vegetative structure of Conocephalum is a thallus which has the appearance of a flattened body of plant tissue.[4][20] The thallus is irregularly branched and relatively large, reaching lengths of roughly 20-24 cm.[4][7] The branching of thalloid liverworts is dichotomous, dividing into two equal parts.[21] The thallus of C. supradecompositum is relatively small, measuring 2-3 cm long.[17] The thallus grows by developing lobes[20] which wither away as the liverwort matures.[22] Plants of C. salebrosum often grow by overlapping lobes, sometimes creating large mats.[4]
Upper surface of the thallus[]
Species of Conocephalum have a thallus that is either dull in appearance, such as C. salebrosum, or distinctly shiny, such as C. conicum.[1][13] The upper surface of the thallus has characteristic polygonal outlines formed by shallow grooves around each air chamber.[7][22] Photosynthetic tissue and chloroplasts are located within the air chambers.[23] In the middle of each air chamber is a white-ringed pore.[7][22] The upper walls of large air chambers are often visible on the surface of the thallus.[22] The air chamber pore remains open, in contrast to the stomata of vascular plants where the pores can open and close.[4]
Underside of the thallus[]
The underside of the thallus has both rhizoids and scales.[7] There are both short and long rhizoids. The short rhizoids are thought to play a role in absorbing water and nutrients. In contrast, the longer rhizoids help anchor the thallus to the underlying substrate. These rhizoids have the appearance of the roots in vascular plants. The rhizoids are single-celled, in contrast to the multicellular rhizoids found in mosses.[20]
Complex oil bodies[]
Liverworts cells often contain complex oil bodies.[3][20][21][24] The oil bodies are intracellular organelles bounded by a single membrane.[23][25] The oil bodies have been known to contain a variety of unique phytochemicals, such as terpenes and flavonoids.[24] The function of oil bodies is still poorly understood.[21][23][25] It has been suggested that oil bodies might function as a deterrent to herbivory or could protect from cold temperatures or harmful ultraviolet radiation.[21][23]
Sporophyte[]
The sporophyte consists of an unbranched stalk called a seta, which bears a terminal spore capsule called a sporangium.[21] The sporangia of Conocephalum are borne beneath stalked gametophytic structures called archegoniophores.[7] In contrast to mosses, the sporophyte matures before the seta elongates.[21][26] Unlike mosses, liverwort sporophytes lack stomata, a columella and peristome teeth.[21][23]
Life cycle[]

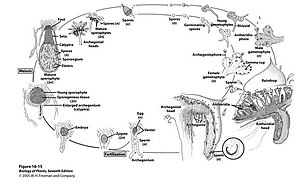
The life cycles of liverworts involves alternating haploid gametophyte and diploid sporophyte generations. The gametophyte generation is dominant, while the diploid sporophyte generation is relatively small and short-lived. The gametophyte produces haploid gametes, egg and sperm, which fuse to form a diploid zygote.The zygote then develops into a sporophyte which ultimately produces haploid spores by meiosis. When spores germinate they first develop into a new free-living individual known as a protonema, before developing into a new thalloid gametophyte.
The sporophyte requires nutrients supplied by the gametophyte to sustain growth and development.
The life cycle of Marchantia liverworts also applies to Conocephalum, with the exception that Conocephalum lacks a stalked antheridiophore.[21]
Reproduction[]


Liverworts reproduce through both sexual and asexual reproduction.[4] In natural populations, the high genetic variation observed suggests that sexual reproduction might dominate.[13] Species of Conocephalum are dioicous,[13] meaning that the male and female reproductive structures are produced on separate plants.[27]
Sexual reproduction[]
Gametophytes produce eggs and sperm in the archegonia and antheridia, respectively.[20] When mature, the antheridia release sperm into the air.[21][28] Fertilization occurs when the sperm reach the egg within the archegonia of a female plant.[22] Once fertilization occurs, the ovule within an archegonium develops into a sporophyte.[20] Mature sporangia on the underside of the receptacle resemble black capsules. These capsules split open to release both spores and elaters,[22] which are dispersed mainly by wind.[20][22] The elaters function to propel spores during dispersal.[26]
Elaters form from an initial mother cell which develops into a diploid cell with spiral thickenings. In contrast, spores develop from an initial diploid mother cell that ultimately forms haploid spores by meiosis.[23] Conocephalum elaters are unique and display a wide range of variability in shape, size and number. Often the abundance of elaters within a capsule are 2-3 times as abundant as spores.[29]
In Conocephalum the male and female reproductive parts are embedded in receptacles on separate plants. On male plants, the receptacle is slightly raised, lacking a stalk, and often circular or oval shaped. Antheridia are embedded in the receptacle[4] and at maturity the sperm is released into the air.[4][21] In contrast, on female plants the receptacles are dome-shaped, with several drooping lobes at the end of an erect stalk. The receptacles are often described as a tiny umbrella, with the archegonia beneath.[4]
Asexual reproduction[]
The production of gemmae is a common method of asexual reproduction in liverworts.[4][26][30] Gemmae are small packets of tissue consisting of haploid cells that are genetically identical with those of the parent plant. They are dispersed by rainfall and ultimately grow into new individuals.[30] In C. conicum, gemmae are located on the lower layers of the thallus and are released as the thallus degrades.[7] The gemmae of C. japonicum are produced from the margins or dorsal surface of the thallus.[31] In contrast, C. salebrosum does not produce gemmae. Vegetative reproduction can occur when a piece of the thallus breaks off and is transported away from the parent plant, for example by water.[4] The individuals resulting from vegetative reproduction are genetically identical to the parent plant and therefore clonal colonies often exist as either all male or all female.[26]
Conocephalum species are perennial, meaning that they can overwinter and produce growth new buds in the spring. These new buds are covered and protected by small scales.[13]
Biochemistry[]
Considerable variations in Conocephalum were identified based on chemical composition. Terpenoids and aromatic compounds are often accumulated within the oil bodies of many liverworts, including Conocephalum. Volatile compounds found in Conocephalum have been used to identify different cryptic species. Considerable variation in these chemical compounds have been identified. Three different groups of C. conicum were identified baed on their unique primary volatile compounds. For example, the compound cubebol, a sesquiterpene alcohol, is characteristic of C. salebrosum.[3] C. supradecompositum has a distinct chemical composition compared to C. conicum as well. It has been demonstrated that the Monoterpenoid content in C. supradecompositum is much less than that of C. conicum.[17]
Associations with other species[]
Fungal interactions[]
Conocephalum can form associations with fungi that are similar to the mycorrhizal associations observed tracheophytes. molecular analyses demonstrated that Conocephalum contained fungal endophytes from the group of fungi known as the Glomeromycota.[32]
C. conicum often colonizes bare soils or rocky substrates and mineral nutrients can therefore be limiting. The fungal endophyte establishes a complex relationship with C. conicum, which is characterized by the formation of arbuscules. These fungi form a highly branched mycelium outside of the plant which then colonize the outside of the rhizoids and pass into the gametophyte. The fungal infection induces proliferation of fungal hypha within the host cells of C. conicum. This association of the fungal hypha with the hosts plastids suggests that photosynthates produced through photosynthesis are likely transferred to the fungus. A similar situation regarding this fungal association has also been identified in the liverwort Pellia epiphylla. Although these associations are common in vascular plants, they have rarely been described in non-vascular plants.[33]
Animal interactions[]
Herbivory[]
Approximately 25 species of moths which are endemic to East Asia associate exclusively with Conocephalum.[13] The larval stage of Epimartyria pardella moths feed on C. conicum.[34][35]
The fungal species Loreleia marchantiae also feeds on C. conicum.
Pathogens[]
The fungal pathogen belonging to the genus Pythium has often been isolated from infected rhizoids and thalli of Conocephalum.[13] Bryoscyphus conocephali is another fungal pathogen that has been associated with C. conicum.[36]
Human applications[]
Conocephalum as a bioindicator for pollution[]
C. conicum has been identified as being tolerant of heavy metals and has therefore been suggested to have a possible role as a bioindicator for pollution. C. conicum takes up ions from both the soil and the atmosphere. Therefore, heavy metals contamination of C. conicum is related not only to air pollution, but environmental contamination from different sources.[37] Recent research has also examined C. conicum as a bioindicator for cadmium pollution. C. conicum was shown to change its biological responses when exposed to cadmium.[38]
Anti-fungal activity[]
C. conicum has been suggested to have a possible role in the management of food borne disease caused by species of Aspergillus fungi. Aspergillus produces highly potent toxins, referred to as aflatoxins, which are harmful to both plants and animals.[24]
Traditional medicine[]
Throughout North America, China and India, liverworts such as Conocephalum have been used for medical purposes. Conocephalum is known to be important to Bhotia, Raji, Tharus and Boxas tribes in Pithoragarh district of Kumaon Himalaya. The extract of C. conicum is also used in treating gallstones.[24]
Photo gallery[]

Conocephalum conicum, the thallus is distinctly shiny

Conocephalum salebrosum, the thallus is dull in appearance

Conocephalum conicum, thallus

Conocephalum conicum, air pores visible on the upper surface of the thallus

Conocephalum salebrosum, thallus cross section showing photosynthetic cells

Conocephalum salebrosum, thallus cross section showing photosynthetic cells

Conocephalum conicum, sporophytes (black) hanging beneath the umbrella-shaped, stalked archegoniophores

Conocephalum salebrosum, sporophytes (black) hanging beneath the umbrella-shaped, stalked archegoniophores

Conocephalum conicum, sporophytes (black) hanging beneath the archegoniophores

Conocephalum conicum, antheridium

Conocephalum salebrosum, ir pore present in the upper surface of the thallus

Conocephalum conicum, air pore present in the upper surface of the thallus












See also[]
- Embryophyte
- Bryophyte
- Marchantiophyta
- Marchantiales
- Conocephalum conicum
- Conocephalum salebrosum
- Conocephalum supradecompositum
- Cryptic species
- Gemmae
- Oil body
- Dioicy
References[]
- ^ a b c d e f g h Szweykowski, J.; Buczkowska, K.; Odrzykoski, I. J. (2005-06-01). "Conocephalum salebrosum (Marchantiopsida, Conocephalaceae) – a new Holarctic liverwort species". Plant Systematics and Evolution. 253 (1): 133–158. doi:10.1007/s00606-005-0301-0. ISSN 1615-6110. S2CID 20076390.
- ^ a b c d AKIYAMA, H. (2022). Morphological and ecological diversification of Conocephalum conicum complex in Japan and Taiwan. Humans Nat, 32, 1-45.
- ^ a b c d e f Ludwiczuk, Agnieszka; Odrzykoski, Ireneusz J.; Asakawa, Yoshinori (2013-11-01). "Identification of cryptic species within liverwort Conocephalum conicum based on the volatile components". Phytochemistry. 95: 234–241. doi:10.1016/j.phytochem.2013.06.011. ISSN 0031-9422. PMID 23835199.
- ^ a b c d e f g h i j k l m n "Minnesota Seasons - snakeskin liverwort". minnesotaseasons.com. Retrieved 2022-02-05.
- ^ a b Zhang, Chi; Chen, Xinlu; Crandall-Stotler, Barbara; Qian, Ping; Köllner, Tobias G.; Guo, Hong; Chen, Feng (2019-08-01). "Biosynthesis of methyl (E)-cinnamate in the liverwort Conocephalum salebrosum and evolution of cinnamic acid methyltransferase". Phytochemistry. 164: 50–59. doi:10.1016/j.phytochem.2019.04.013. ISSN 0031-9422. PMID 31078779. S2CID 153298906.
- ^ a b "Vascular Plants of the Gila Wilderness-- Conocephalum salebrosum". wnmu.edu. Retrieved 2022-02-13.
- ^ a b c d e f g h "Conocephalum conicum | Introduction to Bryophytes". Retrieved 2022-01-24.
- ^ Pryce, Robert J.; Linton, Linda (1974-11-01). "Lunularic acid decarboxylase from the liverwort Conocephalum conicum". Phytochemistry. 13 (11): 2497–2501. doi:10.1016/S0031-9422(00)86926-5. ISSN 0031-9422.
- ^ Webmaster, David Ratz. "Cat-tonque Liverwort - Montana Field Guide". fieldguide.mt.gov. Retrieved 2022-02-05.
- ^ a b c d Stotler, Raymond E.; Crandall-Stotler, Barbara (2017-12-29). "A Synopsis of the Liverwort Flora of North America North of Mexico". Annals of the Missouri Botanical Garden. 102 (4): 574–709. doi:10.3417/2016027. ISSN 0026-6493. S2CID 89829947.
- ^ Ki, H; Nitasaka, Eiji; Odrzykoski, Ireneusz; Yamazak, T (2001-11-01). "Phylogenetic relationships among taxa of the liverwort Conocephalum conicum (Conocephalaceae) revealed by psbA sequence". Genes & Genetic Systems. 76 (5): 279–88. doi:10.1266/ggs.76.279. PMID 11817643.
- ^ a b c d e f g "NatureServe Explorer 2.0". explorer.natureserve.org. Retrieved 2022-01-30.
- ^ a b c d e f g h i j "Bryophyte Ecology Volume 4 ebook | Bryophyte Ecology | Michigan Technological University". digitalcommons.mtu.edu. Retrieved 2022-02-16.
- ^ a b c d Sérgio, C; Vieira, C; Claro, D; Garcia, C (2011-03-01). "Conocephalum salebrosum Szweykowski, Buczkowska & Odrzykoski (Marchantiopsida): modelling the occurrence of a hygrophytic species new to the bryoflora of Portugal, Madeira and Azores". Journal of Bryology. 33 (1): 30–34. doi:10.1179/1743282010Y.0000000004. ISSN 0373-6687. S2CID 85286389.
- ^ "NatureServe Explorer 2.0". explorer.natureserve.org. Retrieved 2022-02-11.
- ^ Odrzykoski, Ireneusz J.; Szweykowski, Jerzy (1991-09-01). "Genetic differentiation without concordant morphological divergence in the thallose liverwortConocephalum conicum". Plant Systematics and Evolution. 178 (3): 135–152. doi:10.1007/BF00937961 (inactive 2022-02-10). ISSN 1615-6110.
{{cite journal}}: CS1 maint: DOI inactive as of February 2022 (link) - ^ a b c Asakawa, Yoshinori; Matsuda, Reiko; Takeda, Reiji (1981-01-01). "Mono- and sesquiterpenoids of Conocephalum supradecompositum". Phytochemistry. 20 (6): 1423–1424. doi:10.1016/0031-9422(81)80055-6. ISSN 0031-9422.
- ^ a b "Species Summary". a100.gov.bc.ca. Retrieved 2022-02-05.
- ^ a b c Cros, R. M., & Buczkowska, K. (2009). Conocephalum salebrosum (Marchantiopsida) new to Spain. Cryptogamie, 30(1), 203.
- ^ a b c d e f g "About Mosses and Liverworts". www.illinoiswildflowers.info. Retrieved 2022-01-21.
- ^ a b c d e f g h i j "Bryophyte Ecology Volume 1 ebook | Bryophyte Ecology | Michigan Technological University". digitalcommons.mtu.edu. Retrieved 2022-02-16.
- ^ a b c d e f g "Snakeskin Liverwort (Conocephalum salebrosum)". www.illinoiswildflowers.info. Retrieved 2022-01-21.
- ^ a b c d e f "Class Marchantiopsida | Introduction to Bryophytes". Retrieved 2022-01-24.
- ^ a b c d Negi, K.; Asthana, A. K.; Chaturvedi, P. (2020-07-01). "GC–MS analysis and antifungal activity of acetone extract of Conocephalum conicum (L) Underw (Liverwort) against aflatoxins producing fungi". South African Journal of Botany. 131: 384–390. doi:10.1016/j.sajb.2020.02.035. ISSN 0254-6299. S2CID 216325490.
- ^ a b He, Xiaolan; Sun, Yu; Zhu, Rui-Liang (2013-09-03). "The Oil Bodies of Liverworts: Unique and Important Organelles in Land Plants". Critical Reviews in Plant Sciences. 32 (5): 293–302. doi:10.1080/07352689.2013.765765. ISSN 0735-2689. S2CID 55444410.
- ^ a b c d "Biology 321 - UBC". www3.botany.ubc.ca. Retrieved 2022-01-30.
- ^ Villarreal, Juan Carlos; Renner, Susanne S. (2013-11-02). "Correlates of monoicy and dioicy in hornworts, the apparent sister group to vascular plants". BMC Evolutionary Biology. 13 (1): 239. doi:10.1186/1471-2148-13-239. ISSN 1471-2148. PMC 4228369. PMID 24180692.
- ^ Shimamura, Masaki; Yamaguchi, Tomio; Deguchi, Hironori (2008). "Airborne sperm of Conocephalum conicum (Conocephalaceae)". Journal of Plant Research. 121 (1): 69–71. doi:10.1007/s10265-007-0128-6. PMID 18058191. S2CID 37902329.
- ^ Kitagawa, Naofumi (1982). "A study of Conocephalum supradecompositum, Marchantiales, Hepaticae". Acta Phytotaxonomica et Geobotanica. 33: 179–189. doi:10.18942/bunruichiri.KJ00001079155.
- ^ a b Smith, AJE (1989). The Liverworts of Britain and Ireland. Cambridge University Press, Cambridge.
- ^ Chaturvedi, SK (2017). "Gemmae in Conocephalum japonicum (Thunb.) Grolle (Conocephalceae) and Anthoceros angustus Steph. (Anthocerotaceae) from Nagaland, North-East India". The International Journal of Plant Reproductive Biology. 9 (2): 187–189. doi:10.14787/ijprb.2017.9.2.187-189 (inactive 2022-02-27).
{{cite journal}}: CS1 maint: DOI inactive as of February 2022 (link) - ^ Ligrone, R.; Carafa, A.; Lumini, E.; Bianciotto, V.; Bonfante, P.; Duckett, J. G. (2007-11-01). "Glomeromycotean associations in liverworts: a molecular, cellular, and taxonomic analysis". American Journal of Botany. 94 (11): 1756–1777. doi:10.3732/ajb.94.11.1756. ISSN 0002-9122. PMID 21636371.
- ^ Ligrone, R.; Lopes, C. (1989). "Cytology and development of a mycorrhiza-like infection in the gametophyte of Conocephalum conicum (L.) Dum. (Marchantiales, Hepatophyta)". New Phytologist. 111 (3): 423–433. doi:10.1111/j.1469-8137.1989.tb00705.x. ISSN 1469-8137. PMID 33874002.
- ^ "Conocephalum conicum | NBN Atlas". species.nbnatlas.org. Retrieved 2022-02-08.
- ^ Davis, Donald R.; Landry, Jean-François (2012-04-19). "A review of the North American genus Epimartyria (Lepidoptera, Micropterigidae) with a discussion of the larval plastron". ZooKeys (183): 37–83. doi:10.3897/zookeys.183.2556. ISSN 1313-2989. PMC 3332027. PMID 22573948.
- ^ "Conocephalum conicum (L.) Dumort. - Encyclopedia of Life". eol.org. Retrieved 2022-02-08.
- ^ Basile, Adriana; Sorbo, Sergio; Conte, Barbara; Cardi, Manuela; Esposito, Sergio (2013-11-01). "Ultrastructural changes and Heat Shock Proteins 70 induced by atmospheric pollution are similar to the effects observed under in vitro heavy metals stress in Conocephalum conicum (Marchantiales – Bryophyta)". Environmental Pollution. 182: 209–216. doi:10.1016/j.envpol.2013.07.014. ISSN 0269-7491. PMID 23933125.
- ^ Maresca, Viviana; Lettieri, Gennaro; Sorbo, Sergio; Piscopo, Marina; Basile, Adriana (2020-09-04). "Biological Responses to Cadmium Stress in Liverwort Conocephalum conicum (Marchantiales)". International Journal of Molecular Sciences. 21 (18): 6485. doi:10.3390/ijms21186485. ISSN 1422-0067. PMC 7555243. PMID 32899890.
- Marchantiales
- Marchantiales genera