Homo antecessor
| Homo antecessor Temporal range: Early Pleistocene,
| |
|---|---|

| |
| The "Boy of Gran Dolina" fossils ATD6-15 (frontal bone) and ATD6-69 (maxilla) at the Museo Arqueológico Nacional de España | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Genus: | Homo |
| Species: | †H. antecessor
|
| Binomial name | |
| †Homo antecessor et al., 1997
| |
Homo antecessor (Latin "pioneer") is an archaic human species recorded in the Spanish Sierra de Atapuerca from 1.2 to 0.8 million years ago during the Early Pleistocene. Populations may have been present elsewhere in Western Europe, and were among the first to colonise that region of the world (hence, the name). The first fossils were found in the Gran Dolina cave in 1994, and the species was formally described in 1997 as the last common ancestor of modern humans and Neanderthals, supplanting the more conventional H. heidelbergensis in this function. H. antecessor has since been reinterpreted as merely an offshoot, though probably one branching off just before the modern human/Neanderthal split.
Despite being so ancient, the face conspicuously parallels the morphology seen in modern humans rather than other archaic humans — namely in its overall flatness as well as the curving of the cheekbone as it merges into the upper jaw — though these elements are known only from a juvenile specimen. Various stature estimates range from 162.3–186.8 cm (5 ft 4 in–6 ft 2 in). H. antecessor may have been broad-chested and rather heavy, much like Neanderthals, though the limbs were proportionally long, a trait more frequent in tropical populations. The kneecaps are thin and have poorly developed tendon attachments. The feet indicate H. antecessor was walking and transmitting body weight differently than modern humans do.
H. antecessor was predominantly manufacturing simple pebbles and flakes out of quartz and chert, though they used a variety of materials. This industry may represent a precursor to the Acheulean industry, which later becomes ubiquitous across Western Eurasia and Africa. Groups may have been dispatching hunting parties, which mainly targeted deer in their savanna and mixed-woodland environment. Many of the H. antecessor specimens were cannibalised, perhaps as a cultural practice or to survive severe famine. There is no evidence they were using fire, and they consequently only inhabited inland Iberia during warm periods, presumably retreating to the coast otherwise.
Taxonomy[]
Research history[]

The Sierra de Atapuerca in northern Spain had long been known to be abundant in fossil remains. The Gran Dolina ("great sinkhole") was first explored for fossils by archaeologist in a short field trip to the region in 1966, where he recovered a few animal fossils and stone tools. He lacked the resources and manpower to continue any further. In 1976, Spanish palaeontologist Trinidad Torres investigated the Gran Dolina for bear fossils (he recovered Ursus remains), but was advised by the Edelweiss Speleological Team to continue at the nearby Sima de los Huesos ("bone pit"). In addition to a wealth of bear fossils, he also recovered archaic human fossils, which prompted a massive exploration of the Sierra de Atapuerca, at first headed by Spanish palaeontologist Emiliano Aguirre but quickly taken over by , Eudald Carbonell, and Juan Luis Arsuaga. They restarted excavation of the Gran Dolina in 1992, and found archaic human remains 2 years later, which in 1997 they formally described as a new species, Homo antecessor.[1] In their original description, Castro and colleagues believed the species was the first human to colonise Europe, hence the name antecessor (Latin for explorer, pioneer, early settler, etc.)[2]

The 25 m (82 ft) of Pleistocene sediments at the Gran Dolina are divided into 11 units, TD1 to TD11 ("trinchera dolina" or "sinkhole trench"). H. antecessor was recovered from TD6, which has consequently become the most well-researched layer of the site. The first field season 1994–1996 excavated a small test pit (to see if the unit warranted further investigation) measuring 6 m2 (65 sq ft). This recovered nearly 100 H. antecessor specimens,[3] the best-preserved being ATD6-15 and ATD6-69 (possibly belonging to the same individual) which most clearly elucidate facial anatomy.[2] In subsequent field seasons from 2003 to 2007, a 13 m2 (140 sq ft) triangular section was excavated, yielding about 70 more specimens.[3] In 2007, an H. antecessor molar was recovered from the nearby Sima del Elefante ("elephant pit") in layer TE9 ("trinchera elefante"), belonging to a 20–25 year old individual. In 2008, the Sima del Elefante yielded an additional mandible fragment, stone flakes, and evidence of butchery.[4]
Additionally, the stone tool assemblage at the Gran Dolina is broadly similar to several other contemporary ones across Western Europe, which may represent the work of the same species, though this is unconfirmable because many of these sites have not produced human fossils.[2] In 2014, 50 footprints dating to between 1.2 million and 800,000 years ago were discovered in Happisburgh, England, which could potentially be attributed to an H. antecessor group given it is the only species identified during that time in Western Europe.[5]
Age and taphonomy[]

The 2003 to 2007 excavations revealed a much more intricate stratigraphy than previously thought, and TD6 was divided into 3 sub-units spanning 13 layers and 9 sedimentary facies. Human presence is recorded in sub-units 1 and 2 and in facies A, D1, and F. Randomly orientated scattered bones were deposited in Facies D1 of layer TD6.2.2 (TD6 sub-unit 2, layer 2) and Facies F of layers TD6.2.2 and TD6.2.3, but in Facies D they seem to have been conspicuously clumped into the northwest area. This might indicate they were dragged into the cave via a debris flow. As for Facies F, which contains the most human remains, they may have been deposited by a floodplain-related geological process inflowing from the main entrance to the northwest, as well as a stronger debris flow from another entrance to the south. Fluvially deposited fossils (dragged in by water) were also recovered from Facies A in layers TD6.2.2, TD6.2.1 and TD6.1.2, indicated by limestone gravel within the size range of the remains. Thus, H. antecessor may not have inhabited the cave, though was at least active nearby. Only 5.6% of the fossils bear any evidence of weathering from open air, roots, and soil, which could mean they were deposited deep into the cave relatively soon after death.[3]
Human occupation seems to have occurred in waves corresponding to timespans featuring a warm, humid savanna habitat (though riversides likely supported woodlands). These conditions were only present during transitions from cool glacial to warm interglacial periods, after the climate warmed and before the forests could expand to dominate the landscape.[7] In 1999, two ungulate teeth from TD6 were dated using uranium–thorium dating to 794 to 668 thousand years ago, further constrained palaeomagnetically to before 780,000 years ago.[8] In 2008, TE9 of the Sima del Elefante was constrained to 1.2–1.1 million years ago using palaeomagnetism and cosmogenic dating.[4] In 2013, TD6 was dated to about 930 to 780 thousand years ago using palaeomagnetism, in addition to uranium–thorium and ESR dating on more teeth.[9] In 2018, ESR dating of the H. antecessor specimen ATD6-92 resulted in an age of 949 to 624 thousand years ago, further constrained palaeomagnetically to before 772,000 years ago.[10]
Until 2013 with the discovery of the 1.4 million year old infant tooth from Barranco León, Orce, Spain, these were the oldest human fossils known from Europe,[11] though human activity on the continent stretches back as early as 1.6 mya in Eastern Europe and Spain indicated by stone tools.[12]
Classification[]
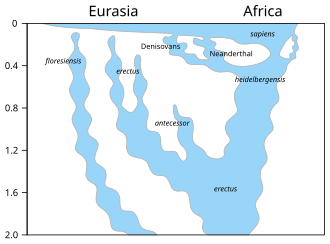
The face of H. antecessor is conspicuously more similar to that of modern humans than to other archaic groups, so in their original description, Castro and colleagues classified it as the last common ancestor between modern humans and Neanderthals, supplanting H. heidelbergensis in this capacity.[2][14][1] The facial anatomy came under close scrutiny in subsequent years.[1]
In 2001, French palaeoanthropologist Jean-Jacques Hublin postulated (without a formal analysis) that the Gran Dolina remains and the contemporaneous Tighennif remains from Algeria (usually classified as Homo ergaster or Homo erectus, originally "Atlantanthropus mauritanicus") represent the same population as 14 of the 15 dental features Castro and colleagues listed for H. antecessor have also been identified in the Middle Pleistocene of North Africa; this would mean H. antecessor is a junior synonym of "Homo mauritanicus", i. e., the Gran Dolina and Tighennif humans should be classified into the latter.[15] In 2003, British palaeoanthropologist Chris Stringer echoed this concern.[16] In 2007, Castro and colleagues formally investigated the matter, and found the Tighennif remains to be much larger than H. antecessor and dentally similar to other African populations. Nonetheless, they still recommended reviving mauritanicus to house all Early Pleistocene North African specimens as "H. ergaster mauritanicus".[17]
In 2007, primatologist Esteban Sarmiento and colleagues questioned the legitimacy of H. antecessor as a separate species because much of the skull anatomy is unknown; H. heidelbergensis is known from roughly the same time and region; and because the type specimen was a child, the supposedly characteristic features could have disappeared with maturity. Such restructuring of the face, they argued, can also be caused by regional climatic adaptation rather than speciation.[18] In 2009, American palaeoanthropologist Richard Klein stated he was skeptical that H. antecessor was ancestral to H. heidelbergensis, interpreting H. antecessor as "an offshoot of H. ergaster [from Africa] that disappeared after a failed attempt to colonize southern Europe".[19] In 2013, anthropologist Sarah Freidline and colleagues suggested the modern humanlike face evolved independently several times among Homo.[20] In 2017, Castro and colleagues conceded that H. antecessor may not be a modern human ancestor, though probably split quite shortly before the modern human/Neanderthal split.[1] In 2020, Dutch-German geneticist Frido Welker and colleagues corroborated this hypothesis by analysing ancient proteins collected from the tooth ATD6-92.[21]
Anatomy[]
Skull[]


The facial anatomy of the 10 to 11.5 year old child ATD6-69 is strikingly similar to modern humans (as well as East Asian Middle Pleistocene archaic humans) as opposed to West Eurasian or African Middle Pleistocene archaic humans or Neanderthals, though African Middle Pleistocene humans (the direct ancestors of modern humans) would later evolve this condition. The most notable traits are a completely flat face and a curved zygomaticoalveolar crest (the bar of bone connecting the cheek to the part of the maxilla which holds the teeth). In 2013, anthropologist Sarah Freidline and colleagues statisticaly determined that these features would not disappear with maturity, H. antecessor suggests the modern human face evolved and disappeared multiple times in the past, which is not unlikely as facial anatomy is strongly influenced by diet and thus the environment.[20] The nasal bones are like that of modern humans.[2] The mandible (lower jaw) is quite gracile unlike most other archaic humans. It exhibits several archaic features, but the shape of the mandibular notch is modern humanlike, and the alveolar part (adjacent to the teeth) is completely vertical. Like many Neanderthals, the medial pterygoid tubercle is large. Unlike most Neanderthals, there is no retromolar space (a large gap between the last molar and the end of the mandible).[22]
The upper incisors are shovel-shaped (the tongue side is distinctly concave), a feature characteristic of other Eurasian human populations. The canines bear the cingulum (towards the base) and the essential ridge (towards the midline) like derived species, but retain the cuspules (small bumps) near the tip and bordering incisor like more archaic species. The upper premolar crowns are rather derived, being nearly symmetrical and bearing a lingual cusp (on the tongue side), and a cingulum and longitudinal grooves on the cheekward side. The upper molars feature several traits typically seen in Neanderthals. The mandibular teeth, on the other hand, are quite archaic. The P3 (the first lower premolar) crowns are strongly asymmetrical and have complex tooth root systems. P3 is smaller than P4 like more derived species, but like other early Homo, M1 (the first lower molar) is smaller than M2 and the cusps of the molar crowns make a Y shape. The distribution of enamel is Neanderthal-like, with thicker layers at the periphery than at the cusps.[22] The thickness of enamel and the proportion of the tooth covered by the gums are conspicuously variable, and may be due to sexual dimorphism, with females having smaller teeth, relatively thicker enamel, and smaller proportion of gum coverage.[23]
The parietal bones (each being one side of the back part of the top of the skull) are flattened, and conjoin at a peak at the midline. This "tent-like" profile is also exhibited in more archaic African H. ergaster and Asian H. erectus. Like H. ergaster, the temporal styloid process just below the ear is fused to the base of the skull. The brow ridges are prominent. The upper margin of the squamous part of temporal bones (on the side of the skull) is convex, like in more derived species.[22]

Torso[]
The notably large adult clavicle specimen ATD6-50, assumed male based on absolute size, was estimated to have stood 162.3–186.8 cm (5 ft 4 in–6 ft 2 in), mean of 174.5 cm (5 ft 9 in), based on the correlation among modern Indian people between clavicle length and stature. An adult radius, ATD6-43, which could be male based on absolute size or female based on gracility, was estimated to have belonged to a 172.5 cm (5 ft 8 in) tall individual based on the average of equations among several modern populations relating radial length to stature. Based on metatarsal (toe bone) length, a male is estimated to have stood 173 cm (5 ft 8 in) and a female 168.9 cm (5 ft 6 in). These are all rather similar values. For comparison, Western European Neanderthal estimates average 165.3 cm (5 ft 5 in), and early European modern humans 178.4 cm (5 ft 10 in).[14] The ankle joint is adapted for handling high stress, which may indicate a heavy, robust body plan, much like Neanderthals.[24] Three of the Happisburgh trackways were laid by individuals measuring approximately 160 cm (5 ft 3 in), 163 cm (5 ft 4 in), and 173 cm (5 ft 8 in), ranging from 48 to 53 kg (106 to 117 lb) based on the relationship between human footprint length and body size. Stature estimates are roughly consistent with those for H. antecessor, H. heidelbergensis, and Neanderthals. The other nine trackways are consistent with individuals under 140 cm (4 ft 7 in), probably children.[5]
Two atlases (the first neck vertebra) are known, which is exceptional as this bone rarely ever fossilises for archaic humans. They are indistinguishable from those of modern humans. For the axis (the second neck vertebra), the angle of the spinous process (jutting out from the vertebra) is about 19°, comparable with Neanderthals and modern humans, diverging from H. ergaster with a low angle of about 8°. The vertebral foramen (which houses the spinal cord) is on the narrow side compared to modern humans. The spine as a whole otherwise aligns with modern humans.[14]
There is one known (and incomplete) clavicle, ATD6-50, which is thick compared to those of modern humans. This may indicate H. antecessor had long and flattish (platycleidic) clavicles like other archaic humans. This would point to a broad chest. The proximal curvature (twisting of the bone on the side nearest the neck) in front-view is on par with that of Neanderthals, but the distal curvature (on the shoulder side) is much more pronounced. The sternum is narrow. The acromion (which extends over the shoulder joint) is small.[14] The shoulder blade is similar to all Homo with a general human body plan, indicating H. antecessor was not as skilled a climber as non-human apes or pre-erectus species, but was capable of efficiently launching projectiles such as stones or spears.[25]
Limbs[]
The incomplete radius, ATD6-43, (a forearm bone) was estimated to have measured 257 mm (10.1 in). It is oddly long and straight for someone from so far north, reminiscent of the proportions seen in early modern humans and many people from tropical populations. This could be explained as retention of the ancestral long-limbed tropical form, as opposed to Neanderthals which evolved shorter limbs. This could also indicate a high brachial index (radial to humeral length ratio). Compared to more recent human species, the cross-section of the radial shaft is rather round and gracile throughout its length. Like archaic humans, the radial neck (near the elbow) is long, giving more leverage to the biceps brachii. Like modern humans and H. heidelbergensis, but unlike Neanderthals and more archaic hominins, the radial tuberosity (a bony knob jutting out just below the radial neck) is anteriorly placed (towards the front side when the arm is facing out).[14]
Like those of other archaic humans, the femur features a developed trochanteric fossa and posterior crest. These traits are highly variable among modern human populations. The two known kneecaps, ATD6-22 and ATD6-56, are subrectangular in shape as opposed to the more common subtriangular, though are rather narrow like those of modern humans. They are quite small and thin, falling at the lower end for modern human females. The apex of the kneecap (the area which does not join to another bone) is not well developed, leaving little attachment for the patellar tendon. The medial (towards the midline) and lateral (towards the sides) facets for the knee joint are roughly the same size in ATD6-56 and the medial is larger in ATD6-22, whereas the lateral is commonly larger in modern humans. The lateral facet encroaches onto a straight flat area as opposed to being limited to a defined vastus notch, an infrequent condition among any human species.[14]
The phalanges and metatarsals of the foot are comparable to those of later humans, but the big toe bone is rather robust, which could be related to how H. antecessor was pushing off the ground. The ankle bone (talus) is exceptionally long and high as well as the facet where it connects with the leg (the trochlea), which may be related to how H. antecessor transmitted body weight. The long trochlea caused a short neck of the talus, which bridges the head of the talus connecting to the toes, and the body of the talus connecting to the leg. This somewhat converges with the condition exhibited in Neanderthals, which is generally explained as a response to a heavy and robust body, to alleviate the consequently higher stress to the articular cartilage in the ankle joint. This would also have permitted greater flexion.[24]
Growth rate[]
In 2010, Castro and colleagues estimated that ATD6-112, represented by a permanent upper and lower first molar, died between 5.3 and 6.6 years of age based on the tooth formation rates in chimpanzees (lower estimate) and modern humans (upper). The molars are hardly worn at all, which means the individual died soon after the tooth erupted, and that the age of first molar eruption occurred at roughly this age. The age is within the range of variation of modern humans, and this developmental landmark can debatably be correlated with life history. If the relation is true, H. antecessor had a prolonged childhood, a characteristic of modern humans in which significant cognitive development takes place.[26]
Pathology[]
The specimen ATD6-69 has an ectopic M3 (upper left third molar), where it erupted improperly, and this caused the impaction of M2, where it was blocked from erupting at all. Ectopia of the wisdom teeth is a common condition in modern humans due to the progressive shortening of the jaws over time, as high as 50% in some populations, but impaction of the neighboring tooth is rare. Impaction can lead to secondary lesions, such as dental cavities, root resorption, or dentigerous or keratocysts.[27]
Culture[]
Technology[]
Stone tools[]



In the lower part of TD6.3 (TD6 subunit 3), 84 lithics were recovered, predominantly small, unmodified quartz pebbles with percussive damage — probably inflicted from pounding items such as bone — as opposed to manufacturing more specialised implements. Though 41% of the section's assemblage consists of flakes, they are rather crude and large — averaging 38 mm × 30 mm × 11 mm (1.50 in × 1.18 in × 0.43 in) — either resulting from rudimentary knapping (stoneworking) skills or difficulty working such poor quality materials. They made use of the unipolar longitudinal method, flaking off only one side of a core, probably to compensate for the lack of pre-planning, opting to knap irregularly shaped and thus poorer-quality pebbles.[6]
Most of the stone tools resided in the lower (older) half of TD6.2, with 831 lithics. The knappers made use of a much more diverse array of materials (though most commonly chert), which indicates they were moving farther out in search of better raw materials. The Sierra de Atapuerca features an abundance and diversity of mineral outcroppings suitable for stone tool manufacturing, in addition to chert and quartz namely quartzite, sandstone, and limestone, which could all be collected within only 3 km (1.9 mi) of the Gran Dolina. They produced far fewer pebbles and spent more time knapping off flakes, but they were not particularly economic with their materials, and about half of the cores could have produced more flakes. They additionally modified irregular blanks into more workable shapes before flaking off pieces. This pre-planning allowed them to use other techniques: the centripetal method (flaking off only the edges of the core) and the bipolar method (laying the core on an anvil and slamming it with a hammerstone). There are 62 flakes measuring below 20 mm (0.79 in) in height, and 28 above 60 mm (2.4 in). There are 3 conspicuously higher quality flakes, thinner and longer than the others, which may have been produced by the same person. There are also retouched tools: notches, spines, denticulates, points, scrapers, and a single chopper. These small retouched tools are rare in the European Early Pleistocene.[6]
TD6.1 yielded 124 lithics, but they are badly preserved as the area was also used by hyenas as a latrine. The layer lacks pebbles and cores, and 44 of the lithics are indeterminate. Flakes are much smaller with an average of 28 mm × 27 mm × 11 mm (1.10 in × 1.06 in × 0.43 in), with 10 measuring below 20 mm (0.79 in), and only 3 exceeding 60 mm (2.4 in). They seem to have been using the same methods as the people who manufactured the TD6.2 tools. They were only retouching larger flakes, the 14 such tools averaging 35 mm × 26 mm × 14 mm (1.38 in × 1.02 in × 0.55 in): 1 marginally retouched flake, 1 notch, 3 spines, 7 denticulate sidescrapers, and 1 denticulate point.[6]
This industry is found elsewhere in Early Pleistocene Spain — notably in Barranc de la Boella and the nearby Galería — distinguished by the preparation and sharpening of cores before flaking, the presence of (crude) bifaces, and some degree of standardisation of tool types. Consequently, they are postulated to represent the ancestor of the Acheulean industry, wherein these and several other techniques would evolve further predominantly in sites across Western Eurasia and Africa.[6]
Fire[]
Only a few charcoal particles have been collected from TD6, which probably originated from a fire well outside the cave. There is no evidence of any fire use or burnt bones (cooking) in the occupation sequences of the Gran Dolina. In other parts of the world, such evidence does not surface in the archaeological record until roughly 400,000 years ago.[28]
These early Europeans probably physiologically withstood the cold, such as by eating a high-protein diet or supporting a heightened metabolism.[28] Despite glacial cycles, the climate was probably similar to that of today's, with the coldest average temperature reaching 2 °C (36 °F) sometime in December and January, and the hottest in July and August 18 °C (64 °F). Freezing temperatures could have been hit from November to March, but the presence of olive and oak suggests subfreezing was an infrequent occurrence.[18] Nonetheless, TD6 occupation sequences seem to have been a few degrees warmer than present-day, and H. antecessor probably migrated from the Mediterranean shore into inland Iberia when colder glacial periods were transitioning to warmer interglacials, and warm grasslands dominated, vacating the region at any other time. They may have followed water bodies while migrating, in the case of Sierra de Atapuerca, most likely the Ebro River.[7] TE9 similarly indicates a generally warm climate, corresponding to the Waalian interglacial.[4] The Happisburgh footprints were lain in estuarine mudflats with open forests dominated by pine, spruce, birch, and in wetter areas alder, with patches of heath and grasslands; the vegetation is consistent with the cooler beginning or end of an interglacial.[5]
Food[]
Sixteen species were recorded from the Gran Dolina, including the bush-antlered deer, the extinct species of fallow deer Dama vallonetensi, the extinct subspecies of red deer Cervus elaphus acoronatus, the extinct bison Bison voigstedtensi, the extinct rhino Stephanorhinus etruscus, the extinct horse Equus stenonis, a monkey, the extinct fox , the extinct bear Ursus dolinensis, the wild boar, a mammoth, the extinct wolf Canis mosbachensis, the spotted hyena, and a lynx. Some specimens of the former 9 species exhibit cut marks consistent with butchery, with about 13% of all Gran Dolina remains bearing some evidence of human modification. Deer are the most commonly butchered animal, with 106 specimens. The inhabitants seem to have carried carcasses back whole when feasible, and only the limbs and skulls of larger quarries. This indicates the Gran Dolina H. antecessor were dispatching hunting parties who killed and hauled back prey to share with the entire group rather than eating their fill beforehand, which evinces social cooperation and division of labour. Less than 5% of all the remains retain animal carnivore damage, in two instances toothmarks overlapping cutmarks, which could indicate animals were sometimes scavenging H. antecessor leftovers.[29]

The Sima del Elefante site records: macaques, boar, bison, fallow deer, the bush-antlered deer, rhinos, E. stenonis, C. mosbachensis, U. dolinensis, the big cat Panthera gombaszoegensis, the lynx Lynx issiodorensis, the extinct fox , and a beaver (in addition to several rats, shrews, and rabbits). The large mammals are most commonly represented by long bones, a few of which are cracked open, presumably to access the bone marrow. Some others bear evidence of percussion and defleshing.[4]
The cool and humid montane environment encouraged the growth of olive, mastic, beech, hazelnut, and chestnut trees, which H. antecessor may have used as food sources, though they become more common in TD7 and TD8 as the interglacial progresses and the environment becomes wetter. In the H. antecessor unit TD6, pollen predominantly derives from juniper and oak. Trees probably grew along rivers and streams, while the rest of the hills and ridges were dominated by grasses.[18]
Cannibalism[]
Eighty adult and child H. antecessor specimens from the Gran Dolina exhibit cut marks, crushing, burning, and other trauma indicative of cannibalism,[14][29] and are the second-most common remains bearing evidence of butchering.[29] Human bodies were efficiently utilised, and may be the reason why most bones are smashed or otherwise badly damaged. There are no complete skull, elements from the face and back of the skull are usually percussed, and the muscle attachments on the face and the base of the skull were cut off. The intense modification of the face was probably to access the brain. The crown of the head was probably struck, resulting in the impact scars on the teeth at the gum line. Several skull fragments exhibit peeling.[30]
The ribs also bear cut marks along the muscle attachments consistent with defleshing, and ATD6-39 has cuts along the length of the rib, which may be related to disembowelment. The nape muscles were sliced off, and the head and neck were probably detached from the body. The vertebrae were often cut, peeled, and percussed. The muscles on all of the clavicles were sawed off to disconnect the shoulder. One radius, ATD6-43, was cut up and peeled. The femur was shattered, probably to extract the bone marrow. The hands and feet variably exhibit percussion, cutting, or peeling, likely a result of dismemberment.[30]
In sum, mainly the meatier areas were prepared, and the rest discarded. This suggests they were butchering humans for nutritional purposes (presumably under dire circumstances), but the face generally exhibits significantly more cutmarks than the faces of animals. When this is seen in prehistoric modern human specimens, it is typically interpreted as evidence of exocannibalism, a form of ritual cannibalism where one eats someone from beyond their social group, such as an enemy from a neighbouring tribe. But, when overviewing the evidence of H. antecessor cannibalism in 1999, Spanish palaeontologist Yolanda Fernandez-Jalvo and colleagues instead ascribed the relative abundance of facial cut marks in the H. antecessor sample to the strongly contrasting structure of the muscle attachments between humans and typical animal prey items (that is, defleshing the human face simply required more cuts, or the butcherers were less familiar with defleshing humans).[30]
Nonetheless, H. antecessor is conspicuously abundant among the butchered, and the assemblage features a dearth of older individuals, composed entirely of young adults and juveniles. In 2010, Carbonell hypothesised that they were practising exocannibalism and hunting down neighbouring tribesmen.[31] In 2019, Spanish palaeoanthropologist Jesús Rodríguez and colleagues argued the demographics can more parsimoniously be explained as the consumption of fellow tribesmen which had already died from natural causes, simply as to not let valuable food go to waste, especially considering the high youth mortality rates in modern hunter-gatherer groups.[32]
See also[]
- Ceprano Man
- Dmanisi hominins
- Early European modern humans
- Happisburgh footprints
- Homo heidelbergensis
- Neanderthal
- Out of Africa I
- Peopling of Europe
- Tautavel Man
References[]
- ^ a b c d de Castro, J.-M. B. (2015). "Homo antecessor: The state of the art eighteen years later". Quaternary International. 433: 22–31. doi:10.1016/j.quaint.2015.03.049.
... a speciation event could have occurred in Africa/Western Eurasia, originating a new Homo clade [...] Homo antecessor [...] could be a side branch of this clade placed at the westernmost region of the Eurasian continent.
- ^ a b c d e Bermudez de Castro, JM; Arsuaga, JL; Carbonell, E; Rosas, A; Martinez, I; Mosquera, M (1997). "A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans" (PDF). Science. 276 (5317): 1392–1395. doi:10.1126/science.276.5317.1392. PMID 9162001. S2CID 31088294. Archived from the original (PDF) on 2020-02-07.
- ^ a b c Campaña, I.; Pérez-González, A.; Benito-Calvo, A.; et al. (2016). "New interpretation of the Gran Dolina-TD6 bearing Homo antecessor deposits through sedimentological analysis". Scientific Reports. 6 (34799): 34799. doi:10.1038/srep34799. PMC 5054435. PMID 27713562.
- ^ a b c d Carbonell, E. (2008). "The first hominin of Europe" (PDF). Nature. 452 (7186): 465–469. Bibcode:2008Natur.452..465C. doi:10.1038/nature06815. hdl:2027.42/62855. PMID 18368116. S2CID 4401629.
- ^ a b c Ashton, N; Lewis, SG; De Groote, I; Duffy, SM; Bates, M; Bates, R; et al. (2014). "Hominin Footprints from Early Pleistocene Deposits at Happisburgh, UK". PLOS ONE. 9 (2): e88329. Bibcode:2014PLoSO...988329A. doi:10.1371/journal.pone.0088329. PMC 3917592. PMID 24516637.
- ^ a b c d e Mosquera, M.; Ollé, A.; Rodríguez-Álvarez, X. P.; Carbonell, E. (2018). "Shedding light on the Early Pleistocene of TD6 (Gran Dolina, Atapuerca, Spain): The technological sequence and occupational inferences". PLOS ONE. 13 (1): 0190889. doi:10.1371/journal.pone.0190889. PMC 5784927. PMID 29370188.
- ^ a b c Blain, H.-L.; Cuenca-Bescós, G.; Burjachs, F.; López-García, J. M.; Lozano-Fernandéz, I.; Rosell, J. (2013). "Early Pleistocene palaeoenvironments at the time of the Homo antecessor settlement in the Gran Dolina cave (Atapuerca, Spain)". Journal of Quaternary Science. 28 (3): 311–319. doi:10.1002/jqs.2622.
- ^ Falguères, Christophe; Bahain, J.; Yokoyama, Y.; Arsuaga, J.; Bermudez de Castro, J.; Carbonell, E.; Bischoff, J.; Dolo, J. (1999). "Earliest humans in Europe: the age of TD6 Gran Dolina, Atapuerca, Spain". Journal of Human Evolution. 37 (3–4): 343–352. doi:10.1006/jhev.1999.0326. PMID 10496991.
- ^ Parés, JM; Arnold, L; Duval, M; Demuro, M; Pérez-Gonzáleza, A; Bermúdez de Castro, JM; Carbonell, E; Arsuagac, JL (2013). "Reassessing the age of Atapuerca-TD6 (Spain): new paleomagnetic results" (PDF). Journal of Archaeological Science. 40 (12): 4586–4595. doi:10.1016/j.jas.2013.06.013.
- ^ Duval, Mathieu; Grün, Rainer; Parés, Josep M.; Martín-Francés, Laura; Campaña, Isidoro; Rosell, Jordi; Shao, Qingfeng; Arsuaga, Juan Luis; Carbonell, Eudald; Bermúdez de Castro, José María (2018). "The first direct ESR dating of a hominin tooth from Atapuerca Gran Dolina TD-6 (Spain) supports the antiquity of Homo antecessor". Quaternary Geochronology. 47: 120–137. doi:10.1016/j.quageo.2018.05.001.
- ^ Toro-Moyano, I; Martínez-Navarro, B; Agustí, J; Souday, C; Bermúdez; de Castro, JM; Martinón-Torres, M; Fajardo, B; Duval, M; Falguères, C; Oms, O; Parés, JM; Anadón, P; Julià, R; García-Aguilar, JM; Moigne, AM; Espigares, MP; Ros-Montoya, S; Palmqvist, P (2013). "The oldest human fossil in Europe, from Orce (Spain)". J Hum Evol. 65 (1): 1–9. doi:10.1016/j.jhevol.2013.01.012. hdl:10261/84112. PMID 23481345.
- ^ Moyano, I. T.; Barsky, D. (2011). "The archaic stone tool industry from Barranco León and Fuente Nueva 3, (Orce, Spain): Evidence of the earliest hominin presence in southern Europe". Quaternary International. 243 (1): 80–91. Bibcode:2011QuInt.243...80M. doi:10.1016/j.quaint.2010.12.011.
- ^ Chris Stringers' hypothesis of the family tree of genus Homo, published in Stringer, C. (2012). "What makes a modern human". Nature 485 (7396): 33–35. doi:10.1038/485033a
- ^ a b c d e f g Carretero, J. M.; Lorenzo, C.; Arsuaga, J. L. (1999). "Axial and appendicular skeleton of Homo antecessor". J. Hum. Evol. 37 (3–4): 459–499. doi:10.1006/jhev.1999.0342. PMID 10496997.
- ^ Hublin, J.-J. (2001). "Northwestern African Middle Pleistocene hominids and their bearing on the emergence of Homo sapiens". In Barham, L.; Robson-Brown, K. (eds.). Human Roots: Africa and Asia in the Middle Pleistocene. Western Academic and Specialist Press. pp. 116–118. ISBN 978-0-9535418-4-3.
- ^ Springer, C. (2003). "Out of Ethiopia". Nature. 423. doi:10.1038/423692a.
- ^ de Castro, J.-M. B.; María-Torres, M.; Gómez-Robles, A.; Prado, L.; Sarmiento, S. (2007). "Comparative analysis of the Gran Dolina-TD6 (Spain) and Tighennif (Algeria) hominin mandibles". Bulletins et Mémoires de la Société of Anthropologie de Paris. 19 (3–4): 149–167. doi:10.4000/bmsap.4623.
- ^ a b c Sarmiento, E. E.; Mowbray, K.; Sawyer, G. J.; Milner, R.; Deak, V.; Tattersall, I. (2007). "Homo antecessor". The Last Human: A Guide to Twenty-two Species of Extinct Humans. Yale University Press. pp. 190–191. ISBN 978-0-300-10047-1.
- ^ Klein, R. (2009). "Hominin Disperals in the Old World". In Scarre, C. (ed.). The Human Past (2nd ed.). p. 108. ISBN 978-0-500-29063-7.
- ^ a b Freidline, S. E.; Gunz, P.; et al. (2013). "Evaluating developmental shape changes in Homo antecessor subadult facial morphology". Journal of Human Evolution. 65 (4): 404–423. doi:10.1016/j.jhevol.2013.07.012. PMID 23998458.
- ^ Welker, Frido; Ramos-Madrigal, Jazmín; Gutenbrunner, Petra; Mackie, Meaghan; Tiwary, Shivani; Rakownikow Jersie-Christensen, Rosa; Chiva, Cristina; Dickinson, Marc R.; Kuhlwilm, Martin; de Manuel, Marc; Gelabert, Pere; Martinón-Torres, María; Margvelashvili, Ann; Arsuaga, Juan Luis; Carbonell, Eudald; Marques-Bonet, Tomas; Penkman, Kirsty; Sabidó, Eduard; Cox, Jürgen; Olsen, Jesper V.; Lordkipanidze, David; Racimo, Fernando; Lalueza-Fox, Carles; Bermúdez de Castro, José María; Willerslev, Eske; Cappellini, Enrico (2020-04-01). "The dental proteome of Homo antecessor". Nature. 580 (7802): 235–238. Bibcode:2020Natur.580..235W. doi:10.1038/s41586-020-2153-8. ISSN 1476-4687. PMC 7582224. PMID 32269345. S2CID 214736611.
- ^ a b c de Castro, J. M. B.; Martinón‐Torres, M.; Arsuaga, J. L.; Carbonell, E. (2017). "Twentieth anniversary of Homo antecessor (1997‐2017): a review". Evolutionary Anthropology. 26 (4): 157–171. doi:10.1002/evan.21540. PMID 28815959. S2CID 11442202.
- ^ García-Campos, C.; Martinén-Torres, M.; Modesto-Mata, M.; Martín-Francés, L.; de Pinillos, M. M.; de Castro, J. M. B. (2021). "Indicators of sexual dimorphism in Homo antecessor permanent canines". Journal of Anthropological Sciences. 99. doi:10.4436/JASS.99001. PMID 33707343.
- ^ a b Pablos, A.; Lorenzo; Martínez, C.; et al. (2012). "New foot remains from the Gran Dolina-TD6 Early Pleistocene site (Sierra de Atapuerca, Burgos, Spain)". Journal of Human Evolution. 63 (4): 610–623. doi:10.1016/j.jhevol.2012.06.008. PMID 22921478.
- ^ García-Martínez, Daniel; Green, David J.; Bermúdez de Castro, José María (2021). "Evolutionary development of the Homo antecessor scapulae (Gran Dolina site, Atapuerca) suggests a modern-like development for Lower Pleistocene Homo". Scientific Reports. 11 (4102): 4102. doi:10.1038/s41598-021-83039-w. PMC 7892855. PMID 33602966.
- ^ de Castro, J.-M. B.; Martinón-Torres, M.; Prado, L.; et al. (2010). "New immature hominin fossil from European Lower Pleistocene shows the earliest evidence of a modern human dental development pattern" (PDF). Proceedings of the National Academy of Sciences. 107 (26): 11739–11744. doi:10.1073/pnas.1006772107. PMC 2900696. PMID 20547843.
- ^ Martín-Francés, L.; Martinón-Torres, M.; de Pinillos, M. M.; Bayle, P.; Fernández-Colón, P.; et al. (2020). "Ectopic maxillary third molar in Early Pleistocene Homo antecessor from Atapuerca-Gran Dolina site (Burgos, Spain)". American Journal of Biological Anthropology. 171 (4): 733–741. doi:10.1002/ajpa.24010.
- ^ a b Roebroekes, W.; Villa, P. (2011). "On the earliest evidence for habitual use of fire in Europe". Proceedings of the National Academy of Sciences. 108 (13): 5209–5214. Bibcode:2011PNAS..108.5209R. doi:10.1073/pnas.1018116108. PMC 3069174. PMID 21402905.
- ^ a b c Saladié, P.; Huguet, R.; et al. (2011). "Carcass transport decisions in Homo antecessor subsistence strategies". Journal of Human Evolution. 61 (4): 425–446. doi:10.1016/j.jhevol.2011.05.012. PMID 21802117.
- ^ a b c Fernández-Jalvo, Y.; Díez, J. C.; Cáceres, I.; Rosell, J. (1999). "Human cannibalism in the Early Pleistocene of Europe (Gran Dolina, Sierra de Atapuerca, Burgos, Spain)". Journal of Human Evolution. 37 (34): 591–622. doi:10.1006/jhev.1999.0324. PMID 10497001. S2CID 25096156.
- ^ Carbonell, E.; Cáceres, I.; Lozano, M.; Saladié, P.; Rosell, J.; Lorenzo, C.; Huguet, R.; Canals, A.; de Castro, J.-M. B. (2010). "Cultural Cannibalism as a Paleoeconomic System in the European Lower Pleistocene". Current Anthropology. 51 (4): 539–549. doi:10.1086/653807. S2CID 1311044.
- ^ Rodríguez, J.; Guillermo, Z.-R.; Ana, M. (2019). "Does optimal foraging theory explain the behavior of the oldest human cannibals?". Journal of Human Evolution. 131: 228–239. doi:10.1016/j.jhevol.2019.03.010. PMID 31182203.
Further reading[]
- de Castro, J.-M. B. (2002). El chico de la Gran Dolina [The Gran Dolina boy] (in Spanish). Crítica. ISBN 978-84-8432-317-4.
External links[]
| Wikimedia Commons has media related to Homo antecessor. |
- Happisburgh Man
- Atapuerca
- Human Timeline (Interactive) – Smithsonian, National Museum of Natural History (August 2016).
Human evolution | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines |
| ||||||||||||||||||||||||||
| Others |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
- 1994 archaeological discoveries
- Early species of Homo
- Lower Paleolithic
- Pleistocene primates
- Prehistoric Spain
- Prehistoric cannibalism
- Fossil taxa described in 1997