Evolution of human intelligence
This article needs additional citations for verification. (April 2021) |
The evolution of human intelligence is closely tied to the evolution of the human brain and to the origin of language. The timeline of human evolution spans approximately 9 million years,[1] from the separation of the genus Pan until the emergence of behavioral modernity by 50,000 years ago. The first 3 million years of this timeline concern Sahelanthropus, the following 2 million concern Australopithecus and the final 2 million span the history of the genus Homo in the Paleolithic era.
Many traits of human intelligence, such as empathy, theory of mind, mourning, ritual, and the use of symbols and tools, are somewhat apparent in great apes, although they are in much less sophisticated forms than what is found in humans like the great ape language.
History[]
Hominidae[]
Hominin timeline | ||||||||||||||||||||
−10 — – −9.5 — – −9 — – −8.5 — – −8 — – −7.5 — – −7 — – −6.5 — – −6 — – −5.5 — – −5 — – −4.5 — – −4 — – −3.5 — – −3 — – −2.5 — – −2 — – −1.5 — – −1 — – −0.5 — – 0 — | Miocene Pliocene Pleistocene Nakalipithecus Oreopithecus Sahelanthropus Orrorin Ardipithecus Australopithecus Homo habilis Homo erectus Homo heidelbergensis Homo sapiens Neanderthals,Denisovans |
| ||||||||||||||||||
(million years ago) | ||||||||||||||||||||
The great apes (hominidae) show some cognitive and empathic abilities. Chimpanzees can make tools and use them to acquire foods and for social displays; they have mildly complex hunting strategies requiring cooperation, influence and rank; they are status conscious, manipulative and capable of deception; they can learn to use symbols and understand aspects of human language including some relational syntax, concepts of number and numerical sequence.[2]
Homininae[]

Around 10 million years ago, the Earth's climate entered a cooler and drier phase, which led eventually to the Quaternary glaciation beginning some 2.6 million years ago. One consequence of this was that the north African tropical forest began to retreat, being replaced first by open grasslands and eventually by desert (the modern Sahara). As their environment changed from continuous forest to patches of forest separated by expanses of grassland, some primates adapted to a partly or fully ground-dwelling life. Here they were exposed to predators, such as the big cats, from whom they had previously been safe.
These environmental pressures caused selection to favor bipedalism: walking on hind legs. This gave the Homininae's eyes greater elevation, the ability to see approaching danger further off, and a more efficient means of locomotion.[citation needed] It also freed the arms from the task of walking and made the hands available for tasks such as gathering food. At some point the bipedal primates developed handedness, giving them the ability to pick up sticks, bones and stones and use them as weapons, or as tools for tasks such as killing smaller animals, cracking nuts, or cutting up carcasses. In other words, these primates developed the use of primitive technology. Bipedal tool-using primates from the subtribe Hominina date back to as far as about 5 to 7 million years ago, such as one of the earliest species, Sahelanthropus tchadensis.
From about 5 million years ago, the hominin brain began to develop rapidly in both size and differentiation of function. There has been a gradual increase in brain volume as humans progressed along the timeline of evolution (see Homininae), starting from about 600 cm3 in Homo habilis up to 1500 cm3 in Homo neanderthalensis. Thus, in general there's a correlation between brain volume and intelligence.[citation needed] However, modern Homo sapiens have a brain volume slightly smaller (1250 cm3) than neanderthals, and the Flores hominids (Homo floresiensis), nicknamed hobbits, had a cranial capacity of about 380 cm3 (considered small for a chimpanzee) about a third of that of H. erectus. It is proposed that they evolved from H. erectus as a case of insular dwarfism. With their three times smaller brain the Flores hominids apparently used fire and made tools as sophisticated as those of their ancestor H.erectus.
Homo[]
Roughly 2.4 million years ago Homo habilis had appeared in East Africa: the first known human species, and the first known to make stone tools, yet the disputed findings of signs of tool use from even earlier ages and from the same vicinity as multiple Australopithecus fossils may put to question how much more intelligent than its predecessors H. habilis was.
The use of tools conferred a crucial evolutionary advantage, and required a larger and more sophisticated brain to co-ordinate the fine hand movements required for this task.[3] Our knowledge of the complexity of behaviour of Homo habilis is not limited to stone culture, they also had habitual therapeutic use of toothpicks.[4] It is speculated that the evolution of a larger brain may have created a problem for early humans, however. A larger brain requires a larger skull, and thus requires the female to have a wider birth canal for the newborn's larger skull to pass through.
The solution to this was to give birth at an early stage of fetal development, before the skull grew too large to pass through the birth canal. This adaptation enabled the human brain to continue to grow, but it imposed a new discipline. It is to be noted that traditional claims about men's and women's gender roles have been challenged in the past years.[5] Regardless, humans' increasingly sedentary lifestyle to protect their more vulnerable offspring led them to grow even more dependent on tool-making to compete with other animals and other humans, and rely less on body size and strength.[citation needed]
About 200,000 years ago Europe and the Middle East were colonized by Neanderthal man, extinct by 39,000 years ago following the appearance of modern humans in the region from 40,000 to 45,000 years ago.
Homo sapiens[]


- Dates approximate, consult articles for details
- (From 2000000 BC till 2013 AD in (partial) exponential notation)
- See also: Java Man (−1.75e+06), Yuanmou Man (−1.75e+06 : -0.73e+06),
- Lantian Man (−1.7e+06), Nanjing Man (- 0.6e+06), Tautavel Man (- 0.5e+06),
- Peking Man (- 0.4e+06), Solo Man (- 0.4e+06), and Peștera cu Oase (- 0.378e+05)
Homo sapiens intelligence[]
The eldest findings of Homo sapiens in Jebel Irhoud, Morocco date back ca. 300,000 years [6][7] Ca. 200,000 year old fossils of Homo sapiens were found in East Africa. It is unclear to what extent these early modern humans had developed language, music, religion, etc.
According to proponents of the Toba catastrophe theory, the climate in non-tropical regions of the earth experienced a sudden freezing about 70,000 years ago, because of a huge explosion of the Toba volcano that filled the atmosphere with volcanic ash for several years. This reduced the human population to less than 10,000 breeding pairs in equatorial Africa, from which all modern humans are descended. Being unprepared for the sudden change in climate, the survivors were those intelligent enough to invent new tools and ways of keeping warm and finding new sources of food (for example, adapting to ocean fishing based on prior fishing skills used in lakes and streams that became frozen).[citation needed]
Around 80,000–100,000 years ago, three main lines of Homo sapiens diverged, bearers of mitochondrial haplogroup L1 (mtDNA) / A (Y-DNA) colonizing Southern Africa (the ancestors of the Khoisan/Capoid peoples), bearers of haplogroup L2 (mtDNA) / B (Y-DNA) settling Central and West Africa (the ancestors of Niger–Congo and Nilo-Saharan speaking peoples), while the bearers of haplogroup L3 remained in East Africa.[citation needed]
The "Great Leap Forward" leading to full behavioral modernity sets in only after this separation. Rapidly increasing sophistication in tool-making and behaviour is apparent from about 80,000 years ago, and the migration out of Africa follows towards the very end of the Middle Paleolithic, some 60,000 years ago. Fully modern behaviour, including figurative art, music, self-ornamentation, trade, burial rites etc. is evident by 30,000 years ago. The oldest unequivocal examples of prehistoric art date to this period, the Aurignacian and the Gravettian periods of prehistoric Europe, such as the Venus figurines and cave painting (Chauvet Cave) and the earliest musical instruments (the bone pipe of Geissenklösterle, Germany, dated to about 36,000 years ago).[8]
The human brain has evolved gradually over the passage of time; a series of incremental changes occurred as a result of external stimuli and conditions. It is crucial to keep in mind that evolution operates within a limited framework at a given point in time. In other words, the adaptations that a species can develop are not infinite and are defined by what has already taken place in the evolutionary timeline of a species. Given the immense anatomical and structural complexity of the brain, its evolution (and the congruent evolution of human intelligence), can only be reorganized in a finite number of ways. The majority of said changes occur either in terms of size or in terms of developmental timeframes.[9]
There have been studies that strongly support the idea that the level of intelligence associated with humans is not unique to our species. Scholars suggest that this could have, in part, been caused by convergent evolution. One common characteristic that is present in species of "high degree intelligence" (i.e. dolphins, great apes, and humans - Homo sapiens) is a brain of enlarged size. Along with this, there is a more developed neocortex, a folding of the cerebral cortex, and von Economo neurons. Said neurons are linked to social intelligence and the ability to gauge what another is thinking or feeling and, interestingly, are also present in bottlenose dolphins.[10] The cerebral cortex is divided into four lobes (frontal, parietal, occipital, and temporal) each with specific functions. The cerebral cortex is significantly larger in humans than in any other animal and is responsible for higher thought processes such as: reasoning, abstract thinking, and decision making.[11]
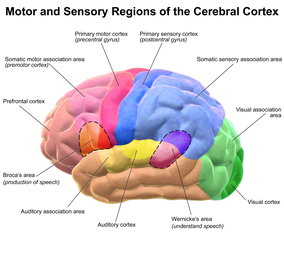
Another characteristic that makes humans special and sets them apart from any other species is our ability to produce and understand complex, syntactic language. The cerebral cortex, particularly in the temporal, parietal, and frontal lobes, are populated with neural circuits dedicated to language. There are two main areas of the brain commonly associated with language, namely: Wernicke's area and Broca's area. The former is responsible for the understanding of speech and the latter for the production of speech. Homologous regions have been found in other species (i.e. Area 44 and 45 have been studied in chimpanzees) but they are not as strongly related to or involved in linguistic activities as in humans.[12]
A big portion of the scholarly literature focus on the evolution, and subsequent influence, of culture. This is in part because the leaps human intelligence has taken are far greater than those that would have resulted if our ancestors had simply responded to their environments, inhabiting them as hunter-gatherers.[13] (Richardson 273).
In short, the immense complexity and marvel of superior human intelligence only emerge inside of a specific culture and history. Selection for cooperation aided our ancestors in surviving harsh ecological conditions and did so by creating a specific type of intelligence. An intelligence that, today, is highly variant from individual to individual.
Models[]
Social brain hypothesis[]
The social brain hypothesis was proposed by British anthropologist Robin Dunbar, who argues that human intelligence did not evolve primarily as a means to solve ecological problems, but rather as a means of surviving and reproducing in large and complex social groups.[14][15] Some of the behaviors associated with living in large groups include reciprocal altruism, deception and coalition formation. These group dynamics relate to Theory of Mind or the ability to understand the thoughts and emotions of others, though Dunbar himself admits in the same book that it is not the flocking itself that causes intelligence to evolve (as shown by ruminants).[14]
Dunbar argues that when the size of a social group increases, the number of different relationships in the group may increase by orders of magnitude. Chimpanzees live in groups of about 50 individuals whereas humans typically have a social circle of about 150 people, which is also the typical size of social communities in small societies and personal social networks;[16] this number is now referred to as Dunbar's number. In addition, there is evidence to suggest that the success of groups is dependent on their size at foundation, with groupings of around 150 being particularly successful, potentially reflecting the fact that communities of this size strike a balance between the minimum size of effective functionality and the maximum size for creating a sense of commitment to the community.[17] According to the social brain hypothesis, when hominids started living in large groups, selection favored greater intelligence. As evidence, Dunbar cites a relationship between neocortex size and group size of various mammals.[14]
Criticism[]
Phylogenetic studies of brain sizes in primates show that while diet predicts primate brain size, sociality does not predict brain size when corrections are made for cases in which diet affects both brain size and sociality. The exceptions to the predictions of the social intelligence hypothesis, which that hypothesis has no predictive model for, are successfully predicted by diets that are either nutritious but scarce or abundant but poor in nutrients.[18] Researchers have found that frugivores tend to exhibit larger brain size than folivores.[19] One potential explanation for this finding is that frugivory requires 'extractive foraging,' or the process of locating and preparing hard-shelled foods, such as nuts, insects, and fruit.[20] Extractive foraging requires higher cognitive processing, which could help explain larger brain size.[20] However, other researchers argue that extractive foraging was not a catalyst in the evolution of primate brain size, demonstrating that some non primates exhibit advanced foraging techniques.[20] Other explanations for the positive correlation between brain size and frugivory highlight how the high-energy, frugivore diet facilitates fetal brain growth and requires spatial mapping to locate the embedded foods.[19]
Meerkats have far more social relationships than their small brain capacity would suggest. Another hypothesis is that it is actually intelligence that causes social relationships to become more complex, because intelligent individuals are more difficult to learn to know.[21]
There are also studies that show that Dunbar's number is not the upper limit of the number of social relationships in humans either.[22][23]
The hypothesis that it is brain capacity that sets the upper limit for the number of social relationships is also contradicted by computer simulations that show simple unintelligent reactions to be sufficient to emulate "ape politics"[24] and by the fact that some social insects such as the paper wasp do have hierarchies in which each individual has its place (as opposed to herding without social structure) and maintains their hierarchies in groups of approximately 80 individuals with their brains smaller than that of any mammal.[25]
Insects provide an opportunity to explore this since they exhibit an unparalleled diversity of social forms to permanent colonies containing many individuals working together as a collective organism and have evolved an impressive range of cognitive skills despite their small nervous systems. [26][27][28] Social insects are shaped by ecology, including their social environment. Studies aimed to correlating brain volume to complexity have failed to identify clear correlations between sociality and cognition because of cases like social insects. In humans, societies are usually held together by the ability of individuals to recognize features indicating group membership. Social insects, likewise, often recognize members of their colony allowing them to defend against competitors. Ants do this by comparing odors which require fine discrimination of multicomponent variable cues. [29] Studies suggest this recognition is achieved through simple cognitive operations that do not involve long-term memory but through sensory adaptation or habituation. [30] In honeybees, their symbolic 'dance' is a form of communication that they use to convey information with the rest of their colony. In an even more impressive social use of their dance language, bees indicate suitable nest locations to a swarm in search of a new home. The swarm builds a consensus from multiple 'opinions' expressed by scouts with different information, to finally agree on a single destination to which the swarm relocates. [31]
Reduction in aggression[]
Another theory that tries to explain the growth of human intelligence is the reduced aggression theory (aka self-domestication theory). According to this strand of thought what led to the evolution of advanced intelligence in Homo sapiens was a drastic reduction of the aggressive drive. This change separated us from other species of monkeys and primates, where this aggressivity is still in plain sight, and eventually lead to the development of quintessential human traits such as empathy, social cognition and culture.[32][33] This theory has received strong support from studies of animal domestication where selective breeding for tameness has, in only a few generations, led to the emergence of impressive "humanlike" abilities. Tamed foxes, for example, exhibit advanced forms of social communication (following pointing gestures), pedomorphic physical features (childlike faces, floppy ears) and even rudimentary forms of theory of mind (eye contact seeking, gaze following).[34][35] Evidence also comes from the field of ethology (which is the study of animal behavior, focused on observing species in their natural habitat rather than in controlled laboratory settings) where it has been found that animals with a gentle and relaxed manner of interacting with each other – like for example stumptailed macaques, orangutans and bonobos – have more advanced socio-cognitive abilities than those found among the more aggressive chimpanzees and baboons.[36] It is hypothesized that these abilities derive from a selection against aggression.[33][37][38][39]
On a mechanistic level these changes are believed to be the result of a systemic downregulation of the sympathetic nervous system (the fight-or-flight reflex). Hence, tamed foxes show a reduced adrenal gland size and have an up to fivefold reduction in both basal and stress-induced blood cortisol levels.[40][41] Similarly, domesticated rats and guinea pigs have both reduced adrenal gland size and reduced blood corticosterone levels.[42][43] It seems as though the neoteny of domesticated animals significantly prolongs the immaturity of their hypothalamic-pituitary-adrenal system (which is otherwise only immature for a short period when they are pups/kittens) and this opens up a larger "socialization window" during which they can learn to interact with their caretakers in a more relaxed way.
This downregulation of sympathetic nervous system reactivity is also believed to be accompanied by a compensatory increase in a number of opposing organs and systems. Although these are not as well specified various candidates for such "organs" have been proposed: the parasympathetic system as a whole, the septal area over the amygdala,[32] the oxytocin system,[44] the endogenous opioids[45] and various forms of quiescent immobilization which antagonize the fight-or-flight reflex.[46][47]
Social exchange theory[]


Other studies suggest that social exchange between individuals is a vital adaptation to the human brain, going as far to say that the human mind could be equipped with a neurocognitive system specialized for reasoning about social change. Social Exchange is a vital adaptation that evolved in social species and has become exceptionally specialized in humans. This adaption will develop by natural selection when two parties can make themselves better off than they were before by exchanging things one party values less for things the other party values for more. However, selection will only pressure social exchange when both parties are receiving mutual benefits from their relative situation; if one party cheats the other by receiving a benefit while the other is harmed, then selection will stop. Consequently, the existence of cheaters—those who fail to deliver fair benefits—threatens the evolution of exchange. Using evolutionary game theory, it has been shown that adaptations for social exchange can be favored and stably maintained by natural selection, but only if they include design features that enable them to detect cheaters, and cause them to channel future exchanges to reciprocators and away from cheaters. Thus, humans use social contracts to lay the benefits and losses each party will be receiving (if you accept benefit B from me, then you must satisfy my requirement R). Humans have evolved an advanced cheater detection system, equipped with proprietary problem-solving strategies that evolved to match the recurrent features of their corresponding problem domains. Not only do humans need to determine that the contract was violated, but also if the violation was intentionally done. Therefore, systems are specialized to detect contract violations that imply intentional cheating.[48]
One problem with the hypothesis that specific punishment for intentional deception could coevolve with intelligence is the fact that selective punishment of individuals with certain characteristics selects against the characteristics in question. For example, if only individuals capable of remembering what they had agreed to were punished for breaking agreements, evolution would have selected against the ability to remember what one had agreed to.[49][50][51] Though this becomes a superficial argument after considering the balancing positive selection for the ability to successfully 'make ones case'. Intelligence predicts the number of arguments one can make when taking either side of a debate. Humans who could get away with behaviours that exploited within and without-group cooperation, getting more while giving less, would overcome this.
In 2004, psychologist Satoshi Kanazawa argued that g was a domain-specific, species-typical, information processing psychological adaptation,[52] and in 2010, Kanazawa argued that g correlated only with performance on evolutionarily unfamiliar rather than evolutionarily familiar problems, proposing what he termed the "Savanna-IQ interaction hypothesis".[53][54] In 2006, Psychological Review published a comment reviewing Kanazawa's 2004 article by psychologists Denny Borsboom and Conor Dolan that argued that Kanazawa's conception of g was empirically unsupported and purely hypothetical and that an evolutionary account of g must address it as a source of individual differences,[55] and in response to Kanazawa's 2010 article, psychologists Scott Barry Kaufman, Colin G. DeYoung, Deirdre Reis, and Jeremy R. Gray gave 112 subjects a 70-item computerized version of the Wason selection task (a logic puzzle) in a social relations context as proposed by Leda Cosmides and John Tooby in The Adapted Mind,[56] and found instead that "performance on non-arbitrary, evolutionarily familiar problems is more strongly related to general intelligence than performance on arbitrary, evolutionarily novel problems".[57][58]
Peter Cathcart Wason originally demonstrated that not even 10% of subjects found the correct solution and his finding was replicated.[59][60] Additionally, psychologists Patricia Cheng, Keith Holyoak, Richard E. Nisbett, and Lindsay M. Oliver demonstrated experimentally that subjects who have completed semester-long college courses in propositional calculus do not perform better on the Wason selection task than subjects who do not complete such college courses.[61] Tooby and Cosmides originally proposed a social relations context for the Wason selection task as part of a larger computational theory of social exchange after they began reviewing the previous experiments about the task beginning in 1983.[56] Despite other experimenters finding that some contexts elicited more correct subject responses than others, no theoretical explanation for differentiating between them was identified until Tooby and Cosmides proposed that disparities in subjects performance on contextualized versus non-contextualized variations of the task was a by-product of a specialized cheater-detection module, and Tooby and Cosmides later noted that whether there are evolved cognitive mechanisms for the content-blind rules of logical inference is disputed.[62][63]
Sexual selection[]
This model, which invokes sexual selection, is proposed by Geoffrey Miller who argues that human intelligence is unnecessarily sophisticated for the needs of hunter-gatherers to survive. He argues that the manifestations of intelligence such as language, music and art did not evolve because of their utilitarian value to the survival of ancient hominids. Rather, intelligence may have been a fitness indicator. Hominids would have been chosen for greater intelligence as an indicator of healthy genes and a Fisherian runaway positive feedback loop of sexual selection would have led to the evolution of human intelligence in a relatively short period.[64] Philosopher Denis Dutton also argued that the human capacity for aesthetics evolved by sexual selection.[65]
Evolutionary biologist George C. Williams and evolutionary medicine researcher Randolph M. Nesse cite evolutionary psychologists John Tooby and Leda Cosmides as referring to the emotions as "Darwinian algorithms of the mind,"[66] while social psychologist David Buss has argued that the sex-specialized differences in the emotion of jealousy are evolutionarily stable strategies for detecting infidelity by a mating partner.[67][68] Citing cross-cultural research conducted by Buss,[69] Miller has argued that if humans prefer altruistic mating partners that would select for altruism directly.[70] Theoretical biologist Mary Jane West-Eberhard and evolutionary medicine researcher Randolph M. Nesse, view sexual selection as a subcategory of social selection,[list 1] with Nesse and anthropologist Christopher Boehm arguing further that altruism in humans held fitness advantages that enabled evolutionarily extraordinary cooperativeness and the human capability of creating culture and collective punishment by bands against bullies, thieves, free-riders, and psychopaths.[list 2]
In many species, only males have impressive secondary sexual characteristics such as ornaments and show-off behavior, but sexual selection is also thought to be able to act on females as well in at least partially monogamous species.[83] With complete monogamy, there is assortative mating for sexually selected traits. This means that less attractive individuals will find other less attractive individuals to mate with. If attractive traits are good fitness indicators, this means that sexual selection increases the genetic load of the offspring of unattractive individuals. Without sexual selection, an unattractive individual might find a superior mate with few deleterious mutations, and have healthy children that are likely to survive. With sexual selection, an unattractive individual is more likely to have access only to an inferior mate who is likely to pass on many deleterious mutations to their joint offspring, who are then less likely to survive.[64]
Sexual selection is often thought to be a likely explanation for other female-specific human traits, for example breasts and buttocks far larger in proportion to total body size than those found in related species of ape.[64] It is often assumed that if breasts and buttocks of such large size were necessary for functions such as suckling infants, they would be found in other species. That human female breasts (typical mammalian breast tissue is small)[84] are found sexually attractive by many men is in agreement with sexual selection acting on human females secondary sexual characteristics.
Sexual selection for intelligence and judging ability can act on indicators of success, such as highly visible displays of wealth. Growing human brains require more nutrition than brains of related species of ape. It is possible that for females to successfully judge male intelligence, they must be intelligent themselves. This could explain why despite the absence of clear differences in intelligence between males and females on average, there are clear differences between male and female propensities to display their intelligence in ostentatious forms.[64]
Critique[]
The sexual selection by the disability principle/fitness display model of the evolution of human intelligence is criticized by certain researchers for issues of timing of the costs relative to reproductive age. While sexually selected ornaments such as peacock feathers and moose antlers develop either during or after puberty, timing their costs to a sexually mature age, human brains expend large amounts of nutrients building myelin and other brain mechanisms for efficient communication between the neurons early in life. These costs early in life build facilitators that reduce the cost of neuron firing later in life, and as a result the peaks of the brain's costs and the peak of the brain's performance are timed on opposite sides of puberty with the costs peaking at a sexually immature age while performance peaks at a sexually mature age. Critical researchers argue the above shows that the cost of intelligence is a signal which reduces the chance of surviving to reproductive age, and does not signal fitness of sexually mature individuals. Since the disability principle is about selection from disabilities in sexually immature individuals, which increases the offspring's chance of survival to reproductive age, disabilities would be selected against and not for by the above mechanism. These critics argue that human intelligence evolved by natural selection citing that unlike sexual selection, natural selection have produced many traits that cost the most nutrients before puberty including immune systems and accumulation and modification for increased toxicity of poisons in the body as a protective measure against predators.[85][86]
Intelligence as a disease-resistance sign[]
The number of people with severe cognitive impairment caused by childhood viral infections like meningitis, protists like Toxoplasma and Plasmodium, and animal parasites like intestinal worms and schistosomes is estimated to be in the hundreds of millions.[87] Even more people with moderate mental damages, such as inability to complete difficult tasks, that are not classified as 'diseases' by medical standards, may still be considered as inferior mates by potential sexual partners.
Thus, widespread, virulent, and archaic infections are greatly involved in natural selection for cognitive abilities. People infected with parasites may have brain damage and obvious maladaptive behavior in addition to visible signs of disease. Smarter people can more skillfully learn to distinguish safe non-polluted water and food from unsafe kinds and learn to distinguish mosquito infested areas from safe areas. Smarter people can more skillfully find and develop safe food sources and living environments. Given this situation, preference for smarter child-bearing/rearing partners increases the chance that their descendants will inherit the best resistance alleles, not only for immune system resistance to disease, but also smarter brains for learning skills in avoiding disease and selecting nutritious food. When people search for mates based on their success, wealth, reputation, disease-free body appearance, or psychological traits such as benevolence or confidence; the effect is to select for superior intelligence that results in superior disease resistance.[citation needed]
Ecological dominance-social competition model[]
A predominant model describing the evolution of human intelligence is ecological dominance-social competition (EDSC),[88] explained by Mark V. Flinn, David C. Geary and Carol V. Ward based mainly on work by Richard D. Alexander. According to the model, human intelligence was able to evolve to significant levels because of the combination of increasing domination over habitat and increasing importance of social interactions. As a result, the primary selective pressure for increasing human intelligence shifted from learning to master the natural world to competition for dominance among members or groups of its own species.
As advancement, survival and reproduction within an increasing complex social structure favored ever more advanced social skills, communication of concepts through increasingly complex language patterns ensued. Since competition had shifted bit by bit from controlling "nature" to influencing other humans, it became of relevance to outmaneuver other members of the group seeking leadership or acceptance, by means of more advanced social skills. A more social and communicative person would be more easily selected.
Intelligence dependent on brain size[]
Human intelligence is developed to an extreme level that is not necessarily adaptive in an evolutionary sense. Firstly, larger-headed babies are more difficult to give birth to and large brains are costly in terms of nutrient and oxygen requirements.[89] Thus the direct adaptive benefit of human intelligence is questionable at least in modern societies, while it is difficult to study in prehistoric societies. Since 2005, scientists have been evaluating genomic data on gene variants thought to influence head size, and have found no evidence that those genes are under strong selective pressure in current human populations.[90] The trait of head size has become generally fixed in modern human beings.[91]
While decreased brain size has strong correlation with lower intelligence in humans, some modern humans have brain sizes as small as with Homo erectus but normal intelligence (based on IQ tests) for modern humans. Increased brain size in humans may allow for greater capacity for specialized expertise.[92]
Expanded cortical regions[]
The two major perspectives on primate brain evolution are the concerted and mosaic approaches.[93] In the concerted evolution approach, cortical expansions in the brain are considered to be a by-product of a larger brain, rather than adaptive potential.[93] Studies have supported the concerted evolution model by finding cortical expansions between macaques and marmosets are comparable to that of humans and macaques.[93] Researchers attribute this result to the constraints on the evolutionary process of increasing brain size.[93] In the mosaic approach, cortical expansions are attributed to their adaptive advantage for the species.[94] Researchers have attributed hominin evolution to mosaic evolution.[94]
Simian primate brain evolution studies show that specific cortical regions associated with high-level cognition have demonstrated the greatest expansion over primate brain evolution.[93] Sensory and motor regions have showcased limited growth.[93] Three regions associated with complex cognition include the frontal lobe, temporal lobe, and the medial wall of the cortex.[93] Studies demonstrate that the enlargement in these regions is disproportionately centered in the temporoparietal junction (TPJ), lateral prefrontal cortex (LPFC), and anterior cingulate cortex (ACC).[93] The TPJ is located in the parietal lobe and is associated with morality, theory of mind, and spatial awareness.[93] Additionally, the Wernicke's area is located in the TPJ.[93] Studies have suggested that the region assists in language production, as well as language processing.[95] The LPFC is commonly associated with planning and working memory functions.[93] The Broca's area, the second major region associated with language processing, is also located in the LPFC.[93] The ACC is associated with detecting errors, monitoring conflict, motor control, and emotion.[93] Specifically, researchers have found that the ACC in humans is disproportionately expanded when compared to the ACC in macaques.[93]
Studies on cortical expansions in the brain have been used to examine the evolutionary basis of neurological disorders, such as Alzheimer's disease.[93] For example, researchers associate the expanded TPJ region with Alzheimer's disease. However, other researchers found no correlation between expanded cortical regions in the human brain and the development of Alzheimer's disease.[96]
Cellular, genetic, and circuitry changes[]
Human brain evolution involves cellular, genetic, and circuitry changes.[97] On a genetic level, humans have a modified FOXP2 gene, which is associated with speech and language development.[98] The human variant of the gene SRGAP2, SRGAP2C, enables greater dendritic spine density which fosters greater neural connections.[99] On a cellular level, studies demonstrate von Economo neurons (VENs) are more prevalent in humans than other primates.[100] Studies show that VENs are associated with empathy, social awareness and self-control.[100] Studies show that the striatum plays a role in understanding reward and pair-bond formation.[101] On a circuitry level, humans exhibit a more complex mirror neuron system, greater connection between the two major language processing areas (Wernicke's area and Broca's area), and a vocal control circuit that connects the motor cortex and brain stem.[97] The mirror neuron system is associated with social cognition, theory of mind, and empathy.[102] Studies have demonstrated the presence of the mirror neuron system in both macaques in humans; However, the mirror neuron system is only activated in macaques when observing transitive movements.[102]
Group selection[]
Group selection theory contends that organism characteristics that provide benefits to a group (clan, tribe, or larger population) can evolve despite individual disadvantages such as those cited above. The group benefits of intelligence (including language, the ability to communicate between individuals, the ability to teach others, and other cooperative aspects) have apparent utility in increasing the survival potential of a group.
In addition, the theory of group selection is inherently tied to Darwin's theory of natural selection. Specifically, that "group-related adaptations must be attributed to the natural selection of alternative groups of individuals and that the natural selection of alternative alleles within populations will be opposed to this development".[103]
Between-group selection can be used to explain the changes and adaptations that arise within a group of individuals. Group-related adaptations and changes are a byproduct of between-group selection as traits or characteristics that prove to be advantageous in relation to another group will become increasingly popular and disseminated within a group. In the end, increasing its overall chance of surviving a competing group.
However, this explanation cannot be applied to humans (and other species, predominantly other mammals) that live in stable, established social groupings. This is because of the social intelligence that functioning within these groups requires from the individual. Humans, while they are not the only ones, possess the cognitive and mental capacity to form systems of personal relationships and ties that extend well beyond those of the nucleus of family. The continuous process of creating, interacting, and adjusting to other individuals is a key component of many species' ecology.
These concepts can be tied to the social brain hypothesis, mentioned above. This hypothesis posits that human cognitive complexity arose as a result of the higher level of social complexity required from living in enlarged groups. These bigger groups entail a greater amount of social relations and interactions thus leading to an expanded quantity of intelligence in humans.[19] However, this hypothesis has been under academic scrutiny in recent years and has been largely disproven. In fact, the size of a species' brain can be much better predicted by diet instead of measures of sociality as noted by the study conducted by DeCasien et al. They found that ecological factors (such as: folivory/frugivory, environment) explain a primate brain size much better than social factors (such as: group size, mating system).[19]
Nutritional status[]
Diets deficient in iron, zinc, protein, iodine, B vitamins, omega 3 fatty acids, magnesium and other nutrients can result in lower intelligence[104][105] either in the mother during pregnancy or in the child during development. While these inputs did not have an effect on the evolution of intelligence they do govern its expression. A higher intelligence could be a signal that an individual comes from and lives in a physical and social environment where nutrition levels are high, whereas a lower intelligence could imply a child, its mother, or both, come from a physical and social environment where nutritional levels are low. Previc emphasizes the contribution of nutritional factors, especially meat and shellfish consumption, to elevations of dopaminergic activity in the brain, which may have been responsible for the evolution of human intelligence since dopamine is crucial to working memory, cognitive shifting, abstract, distant concepts, and other hallmarks of advanced intelligence.[106]
See also[]
- Behavioral modernity
- Denisovan, Dragon Man, Neanderthal, Peking Man
- Human evolution
- Human intelligence
- Primate cognition
- Timeline of human evolution
- Noogenesis
References[]
- ^ Klug WS, Cummings MR, Spencer CA, Palladino MA (2012). Concepts of Genetics (Tenth ed.). Pearson. p. 719. ISBN 978-0-321-75435-6. Lay summary (16 May 2014).
Assuming that chimpanzees and humans last shared a common ancestor about 6.5 million years ago, the tree shows that Neanderthals and humans last shared a common ancestor about 706,000 years ago and that the isolating split between Neanderthals and human populations occurred about 370,000 years ago.
- ^ "Chimpanzee intelligence". Indiana University. 2000-02-23. Retrieved 2008-03-24.
- ^ Ko, Kwang Hyun (2016). "Origins of human intelligence: The chain of tool-making and brain evolution" (PDF). Anthropological Notebooks. 22 (1): 5–22.
- ^ Puech P.-F., Puech S., Cianfarani F., Albertini H. (1989). "Tooth wear and dexterity in Homo erectus", pp. 247–251 in Hominidae. Proceedings of the 2nd International Congress of Human Paleontology, Jaca Book (Milan : Italy)
- ^ Maya, Wei-Haas (November 4, 2020). "Prehistoric female hunter discovery upends gender role assumptions". National Geographic. Retrieved April 7, 2021.
- ^ David Richter et al. (8 June 2017). "The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age". Nature. 546 (7657): 293–296. Bibcode:2017Natur.546..293R. doi:10.1038/nature22335. PMID 28593967. S2CID 205255853.CS1 maint: uses authors parameter (link) "Here we report the ages, determined by thermoluminescence dating, of fire-heated flint artefacts obtained from new excavations at the Middle Stone Age site of Jebel Irhoud, Morocco, which are directly associated with newly discovered remains of H. sapiens8. A weighted average age places these Middle Stone Age artefacts and fossils at 315 ± 34 thousand years ago. Support is obtained through the recalculated uranium series with electron spin resonance date of 286 ± 32 thousand years ago for a tooth from the Irhoud 3 hominin mandible."; Smith TM, Tafforeau P, Reid DJ, et al. (April 2007). "Earliest evidence of modern human life history in North African early Homo sapiens". Proceedings of the National Academy of Sciences of the United States of America. 104 (15): 6128–33. Bibcode:2007PNAS..104.6128S. doi:10.1073/pnas.0700747104. PMC 1828706. PMID 17372199.
- ^ David Richter; et al. (8 June 2017). "The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age". Nature. 546 (7657): 293–296. Bibcode:2017Natur.546..293R. doi:10.1038/nature22335. PMID 28593967. S2CID 205255853. Smith TM, Tafforeau P, Reid DJ, et al. (April 2007). "Earliest evidence of modern human life history in North African early Homo sapiens". Proceedings of the National Academy of Sciences of the United States of America. 104 (15): 6128–33. Bibcode:2007PNAS..104.6128S. doi:10.1073/pnas.0700747104. PMC 1828706. PMID 17372199. Callaway, Ewan (7 June 2017). "Oldest Homo sapiens fossil claim rewrites our species' history". Nature. doi:10.1038/nature.2017.22114. Retrieved 5 July 2017.
- ^ Cross, I., Zubrow, E. and Cowan, F. (2002). "Musical behaviours and the archaeological record: a preliminary study" (PDF). In J. Mathieu (ed.). Experimental Archaeology. 1035. British Archaeological Reports International Series. pp. 25–34. Archived from the original on 2012-06-10.CS1 maint: multiple names: authors list (link) CS1 maint: bot: original URL status unknown (link)
- ^ Iriki A, Sakura O (June 2008). "The neuroscience of primate intellectual evolution: natural selection and passive and intentional niche construction". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 363 (1500): 2229–41. doi:10.1098/rstb.2008.2274. PMC 2394573. PMID 18426757.
- ^ Bearzi M, Stanford CB (2007). "Dolphins and African apes: comparisons of sympatric socio-ecology" (PDF). Contributions to Zoology. 76 (4): 235–254. doi:10.1163/18759866-07604003.
- ^ Brain Facts: A Primer on The Brain and Nervous System. Washington, D.C.: Society for Neuroscience. 2018. p. 51.
- ^ Schenker NM, Hopkins WD, Spocter MA, Garrison AR, Stimpson CD, Erwin JM, Hof PR, Sherwood CC (March 2010). "Broca's area homologue in chimpanzees (Pan troglodytes): probabilistic mapping, asymmetry, and comparison to humans". Cerebral Cortex. 20 (3): 730–42. doi:10.1093/cercor/bhp138. PMC 2820707. PMID 19620620.
- ^ Richardson K (2017-03-21), "Human Intelligence", Genes, Brains, and Human Potential, Columbia University Press, pp. 258–289, doi:10.7312/columbia/9780231178426.003.0009, ISBN 9780231178426
- ^ Jump up to: a b c Dunbar RI (1998). "The social brain hypothesis" (PDF). Evolutionary Anthropology: Issues, News, and Reviews. 6 (5): 178–90. doi:10.1002/(SICI)1520-6505(1998)6:5<178::AID-EVAN5>3.0.CO;2-8.
- ^ Dávid-Barrett T, Dunbar RI (August 2013). "Processing power limits social group size: computational evidence for the cognitive costs of sociality". Proceedings. Biological Sciences. 280 (1765): 20131151. doi:10.1098/rspb.2013.1151. PMC 3712454. PMID 23804623.
- ^ Dunbar, R. I. M. (2014). "The Social Brain: Psychological Underpinnings and Implications for the Structure of Organizations". Current Directions in Psychological Science. 23 (2): 109–114. doi:10.1177/0963721413517118. S2CID 146463887.
- ^ Dunbar, R.I.M. (2014). "The Social Brain: Psychological Underpinnings and Implications for the Structure of Organizations". Current Directions in Psychological Science. 23 (2): 109–114. doi:10.1177/0963721413517118. S2CID 146463887.
- ^ DeCasien, Alex R.; Williams, Scott A.; Higham, James P. (27 March 2017). "Primate brain size is predicted by diet but not sociality". Nature Ecology & Evolution. 1 (5): 112. doi:10.1038/s41559-017-0112. PMID 28812699. S2CID 205564046.
- ^ Jump up to: a b c d DeCasien AR, Williams SA, Higham JP (March 2017). "Primate brain size is predicted by diet but not sociality". Nature Ecology & Evolution. 1 (5): 112. doi:10.1038/s41559-017-0112. PMID 28812699. S2CID 205564046.
- ^ Jump up to: a b c King BJ (August 1986). "Extractive foraging and the evolution of primate intelligence". Human Evolution. 1 (4): 361–372. doi:10.1007/BF02436709. S2CID 83885668.
- ^ Kluger, Jeffrey (August 16, 2010) Inside The Minds of Animals. TIME.
- ^ McCarty, C., Killworth, P.D., Bernard, H.R., Johnsen, E. and Shelley, G. (2000). "Comparing Two Methods for Estimating Network Size" (PDF). Human Organization. 60: 28–39. doi:10.17730/humo.60.1.efx5t9gjtgmga73y. Archived from the original (PDF) on 2012-02-06.CS1 maint: multiple names: authors list (link)
- ^ Bernard, H. Russell; Shelley, Gene Ann; Killworth, Peter (1987). "How Much of a Network does the GSS and RSW Dredge Up?" (PDF). Social Networks. 9: 49–63. doi:10.1016/0378-8733(87)90017-7.
- ^ How the body shapes the way we think: A new view of intelligence, Rolf Pfeifer, Josh Bongard
- ^ Micromonsters, David Attenborough 2013
- ^ Chittka, Lars; Niven, Jeremy (November 2009). "Are Bigger Brains Better?" (PDF). Current Biology. 19 (21): R995–R1008. doi:10.1016/j.cub.2009.08.023. PMID 19922859. S2CID 7247082. Retrieved December 3, 2020.
- ^ Hölldobler, Bert; Wilson, Edward (November 17, 2008). The Superorganism: The Beauty, Elegance, and Strangeness of Insect Societies. ISBN 9780393067040.
- ^ Costa, James (September 30, 2006). The Other Insect Societies. Harvard University Press. ISBN 9780674021631.
- ^ Guerrieri, Fernando J.; Nehring, Volker; Jørgensen, Charlotte G.; Nielsen, John; Galizia, C. Giovanni; d'Ettorre, Patrizia (7 July 2009). "Ants recognize foes and not friends". Proceedings of the Royal Society B: Biological Sciences. 276 (1666): 2461–2468. doi:10.1098/rspb.2008.1860. PMC 2690455. PMID 19364750.
- ^ Bos, Nick; d'Ettorre, Patrizia (2012). "Recognition of Social Identity in Ants". Frontiers in Psychology. 3: 83. doi:10.3389/fpsyg.2012.00083. PMC 3309994. PMID 22461777.
- ^ Seeley, Thomas D. (2010). Honeybee democracy. Princeton, N.J.: Princeton University Press. ISBN 9780691147215.
- ^ Jump up to: a b Eccles, John C. (1989). Evolution of the Brain: Creation of the Self. Foreword by Carl Popper. London: Routledge ISBN 9780415032247.
- ^ Jump up to: a b de Waal, Frans B. M. (1989). Peacemaking among primates. Cambridge, MA: Harvard University Press.ISBN 9780674659216.
- ^ Belyaev, D. K. 1984. "Foxes" pp. 211-214. In Mason I. L. ed., Evolution of Domesticated Animals. Prentice Hall Press.ISBN 0582460468.
- ^ Trut LN, Plyusnina I, Oskina IN (2004). "An experiment on fox domestication and debattable issues of evolution of the dog". Russian Journal of Genetics. 40 (6): 644–655. doi:10.1023/B:RUGE.0000033312.92773.c1. S2CID 19741949.
- ^ "ETHOLOGY". www.indiana.edu. Retrieved 2019-04-10.
- ^ Brothers L, Ring B, Kling A (December 1990). "Response of neurons in the macaque amygdala to complex social stimuli". Behavioural Brain Research. 41 (3): 199–213. doi:10.1016/0166-4328(90)90108-Q. PMID 2288672. S2CID 4023773.
- ^ Brothers L, Ring B (October 1993). "Mesial temporal neurons in the macaque monkey with responses selective for aspects of social stimuli". Behavioural Brain Research. 57 (1): 53–61. doi:10.1016/0166-4328(93)90061-T. PMID 8292255. S2CID 4014644.
- ^ Hare B, Wobber V, Wrangham R (2012). "The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression". Animal Behaviour. 83 (3): 573–585. doi:10.1016/j.anbehav.2011.12.007. S2CID 3415520.
- ^ Osadschuk, L. V. 1997. "Effects of domestication on the adrenal cortisol production of silver foxes during embryonic development ". In In L. N. Trut and L. V. Osadschuk eds., Evolutionary-Genetic and Genetic-Physiological Aspects of Fur Animal Domestication. Oslo: Scientifur.ISSN 0105-2403.
- ^ Trut L, Oskina I, Kharlamova A (March 2009). "Animal evolution during domestication: the domesticated fox as a model". BioEssays. 31 (3): 349–60. doi:10.1002/bies.200800070. PMC 2763232. PMID 19260016.
- ^ Künzl C, Sachser N (February 1999). "The behavioral endocrinology of domestication: A comparison between the domestic guinea pig (Cavia aperea f. porcellus) and its wild ancestor, the cavy (Cavia aperea)". Hormones and Behavior. 35 (1): 28–37. doi:10.1006/hbeh.1998.1493. PMID 10049600. S2CID 33327158.
- ^ Albert FW, Shchepina O, Winter C, Römpler H, Teupser D, Palme R, Ceglarek U, Kratzsch J, Sohr R, Trut LN, Thiery J, Morgenstern R, Plyusnina IZ, Schöneberg T, Pääbo S (March 2008). "Phenotypic differences in behavior, physiology and neurochemistry between rats selected for tameness and for defensive aggression towards humans". Hormones and Behavior. 53 (3): 413–21. doi:10.1016/j.yhbeh.2007.11.010. PMID 18177873. S2CID 4609177.
- ^ Carter CS (2014). "Oxytocin pathways and the evolution of human behavior". Annual Review of Psychology. 65: 17–39. doi:10.1146/annurev-psych-010213-115110. PMID 24050183.
- ^ Nelson EE, Panksepp J (May 1998). "Brain substrates of infant-mother attachment: contributions of opioids, oxytocin, and norepinephrine". Neuroscience and Biobehavioral Reviews. 22 (3): 437–52. doi:10.1016/S0149-7634(97)00052-3. PMID 9579331. S2CID 14345053.
- ^ Porges SW (2003). "Social engagement and attachment: a phylogenetic perspective". Annals of the New York Academy of Sciences. 1008 (1): 31–47. Bibcode:2003NYASA1008...31P. doi:10.1196/annals.1301.004. PMID 14998870. S2CID 1377353.
- ^ Tsoukalas I (2018). "Theory of Mind: Towards an Evolutionary Theory". Evolutionary Psychological Science. 4 (1): 38–66. doi:10.1007/s40806-017-0112-x.Pdf.
- ^ Cosmides L, Barrett HC, Tooby J (May 2010). "Colloquium paper: adaptive specializations, social exchange, and the evolution of human intelligence". Proceedings of the National Academy of Sciences of the United States of America. 107 Suppl 2: 9007–14. Bibcode:2010PNAS..107.9007C. doi:10.1073/pnas.0914623107. PMC 3024027. PMID 20445099.
- ^ Friedrich Nietzsche Beyond Good and Evil: Prelude to a Philosophy of the Future 1886
- ^ Allen, Elizabeth, et al. (1975). "Against 'Sociobiology'". [letter] New York Review of Books 22 (Nov. 13).
- ^ Dawkins, Richard (1979). Twelve misunderstandings of kin selection
- ^ Kanazawa, Satoshi (2004). "General Intelligence as a Domain-Specific Adaptation". Psychological Review. American Psychological Association. 111 (2): 512–523. doi:10.1037/0033-295X.111.2.512. PMID 15065920.
- ^ Kanazawa, Satoshi (2010-02-16). "Why Liberals and Atheists Are More Intelligent". Social Psychology Quarterly. 73 (1): 33–57. CiteSeerX 10.1.1.395.4490. doi:10.1177/0190272510361602. ISSN 0190-2725. S2CID 2642312.
- ^ Kanazawa, Satoshi (May–June 2010). "Evolutionary Psychology and Intelligence Research" (PDF). American Psychologist. 65 (4): 279–289. doi:10.1037/a0019378. PMID 20455621. Retrieved February 16, 2018.
- ^ Borsboom, Denny; Dolan, Conor V. (2006). "Why g is not an adaptation: a comment on Kanazawa (2004)". Psychological Review. 113 (2): 433–437. doi:10.1037/0033-295X.113.2.433. PMID 16637768.
- ^ Jump up to: a b Cosmides, Leda; Tooby, John (1995) [1992]. "3. Cognitive Adaptations for Social Exchange". In Barkow, Jerome H.; Cosmides, Leda; Tooby, John (eds.). The Adapted Mind: Evolutionary Psychology and the Generation of Culture. New York: Oxford University Press. pp. 179–206. ISBN 978-0195101072.
- ^ Kaufman, Scott Barry; DeYoung, Colin G.; Reis, Deidre L.; Gray, Jeremy R. (May–June 2010). "General intelligence predicts reasoning ability even for evolutionarily familiar content" (PDF). Intelligence. 39 (5): 311–322. doi:10.1016/j.intell.2011.05.002. Retrieved February 16, 2018.
- ^ Kaufman, Scott Barry (July 2, 2011). "Is General Intelligence Compatible with Evolutionary Psychology?". Psychology Today. Sussex Publishers. Retrieved February 16, 2018.
- ^ Wason, P. C. (1977). "Self-contradictions". In Johnson-Laird, P. N.; Wason, P. C. (eds.). Thinking: Readings in cognitive science. Cambridge: Cambridge University Press. ISBN 978-0521217569.
- ^ Evans, Jonathan St. B. T.; Newstead, Stephen E.; Byrne, Ruth M. J. (1993). Human Reasoning: The Psychology of Deduction. Psychology Press. ISBN 978-0-86377-313-6.
- ^ Cheng, Patricia; Holyoak, Keith; Nisbett, Richard E.; Oliver, Lindsay M. (1986). "Pragmatic versus syntactic approaches to training deductive reasoning". Cognitive Psychology. Elsevier. 18 (3): 293–328. doi:10.1016/0010-0285(86)90002-2. hdl:2027.42/26121. PMID 3742999. S2CID 10376933. Retrieved April 11, 2020.
- ^ Cosmides, Leda; Tooby, John (2005). "20. Neurocognitive Adaptations Designed for Social Exchange". In Buss, David M. (ed.). The Handbook of Evolutionary Psychology (1st ed.). Hoboken, NJ: Wiley. pp. 600–603. ISBN 978-0471264033.
- ^ Cosmides, Leda; Tooby, John (2016) [2005]. "25. Adaptations for Reasoning About Social Exchange". In Buss, David M. (ed.). The Handbook of Evolutionary Psychology, Volume 2: Integrations (2nd ed.). Hoboken, NJ: Wiley. pp. 625–643. ISBN 978-1118755808.
- ^ Jump up to: a b c d Miller (2008). The Mating Mind. ISBN 978-0-8058-5749-8.
- ^ Dutton, Denis (2009). The Art Instinct: Beauty, Pleasure, and Human Evolution. New York: Bloomsbury Press. pp. 135–163. ISBN 978-1596914018.
- ^ Nesse, Randolph; Williams, George C. (1994). Why We Get Sick: The New Science of Darwinian Medicine. New York: Vintage Books. pp. 209–212. ISBN 978-0679746744.
- ^ Buss, David M. (2016) [1994]. The Evolution of Desire: Strategies of Human Mating (3rd ed.). New York: Basic Books. pp. 189–221. ISBN 978-0465097760.
- ^ Buss, David M. (2011) [2000]. The Dangerous Passion: Why Jealousy Is as Necessary as Love and Sex (2nd ed.). New York: Free Press. ISBN 978-1451673135.
- ^ Buss, David M. (1989). "Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures". Behavioral and Brain Sciences. Cambridge University Press. 12 (1): 1–49. doi:10.1017/S0140525X00023992.
- ^ Miller, Geoffrey F. (2000). The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature (1st ed.). New York: Doubleday. pp. 292–340. ISBN 978-0385495165.
- ^ West-Eberhard, Mary Jane (1975). "The Evolution of Social Behavior by Kin Selection". The Quarterly Review of Biology. University of Chicago Press. 50 (1): 1–33. doi:10.1086/408298. JSTOR 2821184. S2CID 14459515.
- ^ West-Eberhard, Mary Jane (1979). "Sexual Selection, Social Competition, and Evolution". Proceedings of the American Philosophical Society. American Philosophical Society. 123 (4): 222–34. JSTOR 986582.
- ^ West-Eberhard, Mary Jane (1983). "Sexual Selection, Social Competition, and Speciation". Quarterly Review of Biology. University of Chicago Press. 58 (2): 155–183. doi:10.1086/413215. JSTOR 2828804. S2CID 54711556.
- ^ Lyon, Bruce E.; Montgomerie, Robert (2012). "Sexual selection is a form of social selection". Philosophical Transactions of the Royal Society B. London, UK: Royal Society. 367 (1600): 2266–2273. doi:10.1098/rstb.2012.0012. PMC 3391428. PMID 22777015.
- ^ West-Eberhard, Mary Jane (2014). "Darwin's forgotten idea: the social essence of sexual selection". Neuroscience & Biobehavioral Reviews. Elsevier. 46 (4): 501–508. doi:10.1016/j.neubiorev.2014.06.015. PMID 25003806. S2CID 1604935.
- ^ Nesse, Randolph (2019). Good Reasons for Bad Feelings: Insights from the Frontier of Evolutionary Psychiatry. Dutton. pp. 172–176. ISBN 978-1101985663.
- ^ Boehm, Christopher (1999). "The Natural Selection of Altruistic Traits" (PDF). Human Nature. Springer Science+Business Media. 10 (3): 205–252. doi:10.1007/s12110-999-1003-z. PMID 26196335. S2CID 207392341. Retrieved July 7, 2021.
- ^ Boehm, Christopher (2001) [1999]. Hierarchy in the Forest: The Evolution of Egalitarian Behavior (Revised ed.). Cambridge, MA: Harvard University Press. ISBN 978-0674006911.
- ^ Nesse, Randolph M. (2007). "Runaway social selection for displays of partner value and altruism". Biological Theory. Springer Science+Business Media. 2 (2): 143–55. doi:10.1162/biot.2007.2.2.143. S2CID 195097363.
- ^ Nesse, Randolph M. (2009). "10. Social Selection and the Origins of Culture". In Schaller, Mark; Heine, Steven J.; Norenzayan, Ara; Yamagishi, Toshio; Kameda, Tatsuya (eds.). Evolution, Culture, and the Human Mind. Philadelphia: Taylor & Francis. pp. 137–50. ISBN 978-0805859119.
- ^ Boehm, Christopher (2012). Moral Origins: Social Selection and the Evolution of Virtue, Altruism, and Shame. New York: Basic Books. ISBN 978-0465020485.
- ^ Boehm, Christopher (2014). "The moral consequences of social selection". Behaviour. Brill Publishers. 151 (2–3): 167–183. doi:10.1163/1568539X-00003143. Retrieved July 7, 2021.
- ^ Jones, Ian L.; Hunter, Fiona M. (1993). "Mutual sexual selection in a monogamous seabird". Nature. 362 (6417): 238–239. Bibcode:1993Natur.362..238J. doi:10.1038/362238a0. S2CID 4254675.
- ^ "Heredity and Society". Author Ian Porter. 1972. Page 51.
- ^ The Evolution of Intelligence; Robert J. Sternberg, James C. Kaufman, 2013
- ^ The First Idea: How Symbols, Language, and Intelligence Evolved from Our Primate Ancestors to Modern Humans; Stanley I. Greenspan, Stuart Shanker, 2009
- ^ Olness K (April 2003). "Effects on brain development leading to cognitive impairment: a worldwide epidemic". Journal of Developmental and Behavioral Pediatrics. 24 (2): 120–30. doi:10.1097/00004703-200304000-00009. PMID 12692458. S2CID 31999992.
- ^ Flinn, M. V.; Geary, D. C.; Ward, C. V. (2005). "Ecological dominance, social competition, and coalitionary arms races: Why humans evolved extraordinary intelligence" (PDF). Evolution and Human Behavior. 26 (1): 10–46. doi:10.1016/j.evolhumbehav.2004.08.005.
- ^ Isler K, van Schaik CP (December 2006). "Metabolic costs of brain size evolution". Biology Letters. 2 (4): 557–60. doi:10.1098/rsbl.2006.0538. PMC 1834002. PMID 17148287.
- ^ Ossorio PN (2011). "Chapter 9: Myth and Mystification: The Science of Race and IQ". In Krimsky S, Sloan K (eds.). Race and the Genetic Revolution: Science, Myth, and Culture. Columbia University Press. p. 189. ISBN 978-0-231-52769-9. Retrieved 31 August 2013. Lay summary (31 August 2013).
Since 2005, other researchers have evaluated the same data on MCHPI and ASPM, plus some additional data, and concluded that there is no evidence that these genes have been under natural selection in modern humans. These reanalyses undercut the idea that the particular variants found at high frequency among people of European descent somehow made Europeans better adapted for modern civilization. Additional studies have discovered that the MCHPI and ASPM variants reported in the 2005 papers do not correlate with larger (or smaller) than average head size. The genes were originally described as having to do with head size because some variants of these genes can cause microcephaly (extremely small heads that lack major portions of the brain). However, those microcephaly-causing variants were not included in the studies published in 2005. Finally, several research groups have tried and failed to show any correlation between the variants described in the 2005 papers and IQ reading abilities, or verbal abilities. (citing "Comment on 'Ongoing Adaptive Evolution of ASPM, a Brain Size Determinant in Homo Sapiens' and 'Microcephalin, a Gene Regulating Brain Size Continues to Evolve Adaptively in Humans," Science 313 (2006): 172 (a); F.Yu, S.R.Hill, S.F.Schaffner, et al., "Comment on 'Ongoing Adaptive Evolution of ASPM, a Brain Size Determinant in Homo Sapiens,'" Science 316 (2007): 370 (b). )
- ^ Klug WS, Cummings MR, Spencer CA, Palladino MA (2012). Concepts of Genetics (Tenth ed.). Pearson. p. 669. ISBN 978-0-321-75435-6. Lay summary (16 May 2014).
h2 values vary, but heritability tends to be low for quantitative traits that are essential to an organism's survival. Remember, this does not indicate the absence of a genetic contribution to the observed phenotypes for such traits. Instead, the low h2 values show that natural selection has already largely optimized the genetic component of these traits during evolution.
- ^ Skoyles JR (1999). "Human evolution expanded brains to increase expertise capacity, not IQ" (PDF). Psycoloquy. 10 (2).
- ^ Jump up to: a b c d e f g h i j k l m n o Evolution of nervous systems. Kaas, Jon H. (Second ed.). Kidlington, Oxford, United Kingdom. 2016-11-23. ISBN 9780128040966. OCLC 964620094.CS1 maint: others (link)
- ^ Jump up to: a b Foley RA (July 2016). "Mosaic evolution and the pattern of transitions in the hominin lineage". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1698): 20150244. doi:10.1098/rstb.2015.0244. PMC 4920300. PMID 27298474.
- ^ Binder JR (December 2015). "The Wernicke area: Modern evidence and a reinterpretation". Neurology. 85 (24): 2170–5. doi:10.1212/WNL.0000000000002219. PMC 4691684. PMID 26567270.
- ^ Fjell AM, Amlien IK, Sneve MH, Grydeland H, Tamnes CK, Chaplin TA, Rosa MG, Walhovd KB (September 2015). "The Roots of Alzheimer's Disease: Are High-Expanding Cortical Areas Preferentially Targeted?†". Cerebral Cortex. 25 (9): 2556–65. doi:10.1093/cercor/bhu055. PMC 6276920. PMID 24658616.
- ^ Jump up to: a b Sherwood CC, Schumacher M (September 2018). "What Makes the Human Brain Special". Scientific American.
- ^ "FOXP2 gene". Genetics Home Reference. Retrieved 2019-04-11.
- ^ Charrier C, Joshi K, Coutinho-Budd J, Kim JE, Lambert N, de Marchena J, et al. (May 2012). "Inhibition of SRGAP2 function by its human-specific paralogs induces neoteny during spine maturation". Cell. 149 (4): 923–35. doi:10.1016/j.cell.2012.03.034. PMC 3357949. PMID 22559944.
- ^ Jump up to: a b Allman JM, Tetreault NA, Hakeem AY, Manaye KF, Semendeferi K, Erwin JM, Park S, Goubert V, Hof PR (April 2011). "The von Economo neurons in the frontoinsular and anterior cingulate cortex". Annals of the New York Academy of Sciences. 1225 (1): 59–71. Bibcode:2011NYASA1225...59A. doi:10.1111/j.1749-6632.2011.06011.x. PMC 3140770. PMID 21534993.
- ^ Báez-Mendoza R, Schultz W (December 2013). "The role of the striatum in social behavior". Frontiers in Neuroscience. 7: 233. doi:10.3389/fnins.2013.00233. PMC 3857563. PMID 24339801.
- ^ Jump up to: a b Rajmohan V, Mohandas E (January 2007). "Mirror neuron system". Indian Journal of Psychiatry. 49 (1): 66–9. doi:10.4103/0019-5545.31522. PMC 2900004. PMID 20640069.
- ^ Williams, George C and Dawkins, R. "Group Selection." Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton University Press, Princeton; Oxford, 2019, pp. 92–124.
- ^ Melville K (22 November 2004). "Poor Nutrition Leads to Low IQ". scienceagogo.com.
- ^ Hoffer A (1989). "Vitamin and Mineral Supplements Increase Intelligence". Nutrition Health Review. 52.
- ^ Previc FH (2009). The dopaminergic mind in human evolution and history. Cambridge, England: Cambridge University Press. ISBN 978-0-521-51699-0.
- Bundled references
Further reading[]
- Byrne RW (1995). The Thinking Ape: Evolutionary origins of intelligence. Oxford, England: Oxford University Press. ISBN 978-0-19-852188-4.
- Greenspan SI, Shanker S (2004). The First Idea: How symbols, language, and intelligence evolved from our early primate ancestors to modern humans. Cambridge, Mass.: Da Capo Press. ISBN 978-0-7382-0680-6.
- Skoyles J, Sagan D (2002). Up from Dragons: The evolution of human intelligence. New York: McGraw-Hill. ISBN 978-0-07-137825-3.
- Tobias, Phillip V. (1971). The Brain in Hominid Evolution. New York: Columbia University Press. ISBN 978-0-231-03518-7.
- Roth G, Dicke U (May 2005). "Evolution of the brain and intelligence". Trends in Cognitive Sciences. 9 (5): 250–7. doi:10.1016/j.tics.2005.03.005. PMID 15866152. S2CID 14758763.
- Todorov A, Fiske S, Prentice DA (11 February 2011). Social Neuroscience: Toward Understanding the Underpinnings of the Social Mind. Oxford University Press, USA. ISBN 978-0-19-531687-2.
| show Evolutionary psychology |
|---|
| show Human evolution |
|---|
| show Evolutionary biology |
|---|

- Human evolution
- Intelligence