Gigantoraptor
| Gigantoraptor Temporal range: Late Cretaceous,
~96 Ma PreꞒ
Ꞓ
O
S
D
C
P
T
J
K
Pg
N
↓ | |
|---|---|

| |
| Mounted reconstructed skeleton of Gigantoraptor in Japan | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Superfamily: | †Caenagnathoidea |
| Family: | †Caenagnathidae |
| Genus: | †Gigantoraptor Xu et al., 2007 |
| Type species | |
| †Gigantoraptor erlianensis Xu et al., 2007
| |
Gigantoraptor (meaning "giant seizer") is a genus of large oviraptorosaur dinosaur that lived in Asia during the Late Cretaceous period. It is known from the Iren Dabasu Formation of Inner Mongolia, where the first remains were found in 2005.
Gigantoraptor reached massive sizes; it is estimated to have cover 8–8.9 m (26–29 ft) in length, and to have weighed about 2 to 2.7 t (2,000 to 2,700 kg). It had an extensively pneumatized vertebral column and elongated arms and legs. Both femur and tibia measured over 1 m (3.3 ft) in length, an unusual trait among giant theropods. The lower jaws were toothless and ended in a keratinous beak, as seen in other oviraptorosaurs. Though several oviraptorosaur species are known to have developed a full coat of feathers, Gigantoraptor, due to its size, could have lost some of this integument.
The genus is classified as an oviraptorosaurian dinosaur, a group of generally small feathered animals. Though it was originally found to represent a basal oviraptorid, subsequent analyses have shown it to be a caenagnathid. It was a giant, ground-dwelling bipedal omnivore with a shearing bite as indicated by the preserved mandible. The shape of its beak indicates a generalist diet with a potentially occasional carnivory. The holotype—and only known specimen—has been determined to represent a young adult that died at the age of 11, and it reached a young adulthood around 7 years of life. Such development indicates an accelerated growth compared to other larger theropods. The discovery and examination of large oviraptorosaur eggs, Macroelongatoolithus, indicates that large species such as Gigantoraptor built their nests with the center lacking eggs in order to avoid crushing.
History of discovery[]
In a quarry at , Iren Dabasu Formation, Erlian basin, Sonid Left Banner (Inner Mongolia), numerous remains of the sauropod Sonidosaurus have been uncovered since 2001. Chinese paleontologist Xu Xing was asked to reenact the discovery of Sonidosaurus in April 2005 for a Japanese documentary. Xu obliged them by digging out a thighbone. As he wiped the bone clean, he suddenly realized it was not from a sauropod, but from an unidentified theropod in the size class of Albertosaurus. He then stopped the filming to secure the serendipitous find. This way, the discovery of the Gigantoraptor holotype fossil was documented on film.[1][2]
That holotype, V0011, consists of the incomplete and disassociated remains of a single adult individual, preserving a nearly complete mandible, a partial isolated cervical vertebra, dorsal vertebrae, caudal vertebrae, right scapula, right humerus, right radius and ulna, nearly complete right manus, partial ilium with a nearly complete pubis and hindlimbs, including both femur, tibia and fibula with a very complete pes.[1]
In 2007, the type species Gigantoraptor erlianensis was named and described by Xu, , , Zhao Xijin and Tan Lin. The generic name, Gigantoraptor, is derived from the Latin gigas, gigantis, meaning "giant" and raptor, meaning "seizer". The specific name, erlianensis, refers to the .[1] The holotype is currently housed at the Longhao Institute of Geology and Paleontology, Inner Mongolia.[3]
Description[]
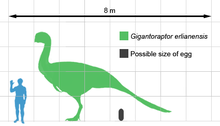
Gigantoraptor is the largest known oviraptorosaur for which skeletal material is available. Approximately 3 times as long and 35 times more heavy than the largest earlier discovered oviraptorosaur Citipati, the holotype of Gigantoraptor has been estimated at 8 m (26 ft) long with a height of 3.5 m (11 ft) at the hips, and a ponderous weight of 1.4 t (1,400 kg).[1] Other estimates have suggested a length of 8.9 m (29 ft) and a mass ranging between 2 to 2.7 t (2,000 to 2,700 kg).[4][5][6]
Oviraptorosaurs are fairly known to have developed feathers, as seen on feathered specimens of Caudipteryx and the pygostyle of several oviraptorids.[1][7] As an oviraptorosaur, Gigantoraptor was likely feathered as well, however, given its massive size it may have been partially naked.[1]
Skull[]



As preserved, the total length of the lower jaws is 46 cm (460 mm). This element fused into a broad shovel-like shape, indicating that the unknown skull was over half a metre long and toothless, likely equipped with a rhamphotheca (horny beak). Compared to the right side, the left side of the jaws is well-preserved with an almost intact shape. The dentary bones are very deep and toothless, and have a pair of sharply-developed shelves on the top edges. These structures are relatively flat. The anterior end of the dentary is rounded, more similar to caenagnathids and different from the chin-shaped one of some oviraptorids. From a top view, the symphyseal (bone union) region at the front of the dentary is U-shaped. Along the lateral surfaces of this bone, some nutrient foramina can be found, which likely supported a rhamphotheca.[3]
Both angular and articular-surangular-coronoid (ASC) complex are comparatively smaller than the dentary. The angular is firmly fused to the ASC complex and gives form to the lower margin of the external mandibular fenestra. The angular has a bowed shape and is mainly forwards extended with a large lateral depression that joints with the posterior lower process (bony projection/extension) of the dentary. The ASC complex is composed by the fusion of the articular and surangular bones with the coronoid process of the mandible. This complex extends forwards to articulate with the posterior upper process of the dentary and gives form to the upper margin of the external mandibular fenestra. The coronoid process prominence is low and distinctively rounded. On the posterior end of the mandibles the articular glenoid can be located. It is relatively large, convex, and bowl-shaped.[3]
Postcranial skeleton[]

The anterior caudal vertebrae have very long neural spines and are heavily pneumaticised with deep pleurocoels. The middle section of the relatively short tail is somewhat stiffened by long prezygapophyses. The caudal vertebrae are heavily pneumatized with a spongeous-like bone filling. The forelimbs are rather long because of an elongated humerus (73.5 cm (735 mm) long) and slender manus. The humerus is bowed outwards to an exceptionally large extent and has a very rounded head, similar to some birds. The first metacarpal is very short and carries a strongly diverging thumb. The hindlimbs are well-developed and highly elongated: the femur is slender with a distinct head and neck, measuring 1.10 m (110 cm); the tibia has similar top and bottom borders and measures 1.10 m (110 cm) long; and the metatarsus is 58.3 cm (583 mm) long with the metatarsal III being the largest. The pes is robust with large and strongly curved pedal unguals.[1][8]
Distinguishing characteristics[]

According to Xu et al. 2007, Gigantoraptor can be recognised by the following traits: reduced mandible with 45% less length compared to the femur; anterior caudal vertebrae with elongated neural spines and posteriorly located stocky, rod-like transverse processes; dentary with elongated postero-ventral process reaching to the level of the glenoid; reduced posteriorly tapered retro articular process much deeper than its wide; anterior and middle caudal vertebrae with a large pneumatic opening on the ventral surface; dentary with two fossa on the lateral surface and close to the external mandibular fenestra; anterior caudal centrum with postero-ventral margin extending ventrally; advanced laminal system of the anterior caudal vertebrae; vertical prezygapophyseal articular facets located proximal to the distal end of the process in the middle caudal vertebrae; scapula with prominent convexity ventral to the acromion process on the lateral surface; reduced calcaneum obscured from anteriorview by the expanded astragalus; distal tarsal IV with proximal projection on the lateral margin; anterior caudal vertebrae composed of opisthocoelous, amphicoelous and procoelous; pleurocoels present on most caudal vertebrae; anterior caudal vertebrae centrum with a pair of vertically arranged pneumatic openings on the lateral surface; bowed humerus with a prominent, spherical top and a curved delto-pectoral crest; proximal section of the humerus with a centrally constricted thick ridge running along the posterior margin; sub-circular, concave proximal articular surface present in the straight ulna; metatarsal III with ginglymoid distal end; constricted proximal articular surface and two lateral splines present in the pedal unguals; radius featuring a sub-spherical distal end; convex medial margin of the proximal end and a medial condyle three times elongated and extending further more distally than the lateral one on the distal end of the metacarpal I; prominent dorso-lateral process on the proximal end and a longitudinal spline on the ventral margin of the proximal third of the end in the metacarpal II; narrow spline-medial to the trochanteric head extending down to the posterior margin of the femoral end, and a patellar spline present on the anterior surface of the distal end; manual unguals with triangular lateral splines; pubis laterally compressed; femur with straight end; neck tightened between the postero-medially oriented spherical femoral top and the anteroposteriorly expanded trochanteric crest that is more stocky and higher anteriorly than posteriorly.[1]
Classification[]
In 2007, Xu et al. assigned Gigantoraptor to the Oviraptoridae, in a basal position. The anatomy of Gigantoraptor includes the diagnostic features of the Oviraptorosaurs. However, it also includes several features found in more derived eumaniraptoran dinosaurs, such as a forelimb/hindlimb ratio of 60%, a lack of expansion of the distal scapula and the lack of a fourth trochanter on the femur. Despite its size, Gigantoraptor would thus have been more bird-like than its smaller oviraptorosaurian relatives.[1]
In 2010, a second analysis of Gigantoraptor relationships found it to be a member of the Caenagnathidae rather than an oviraptorid.[9] Phylogenetic analysis conducted by Lamanna et al. (2014), supported that Gigantoraptor was a basal caenagnathid.[10]

The cladogram below follows the phylogenetic analysis conducted by Funston & Currie in 2016, which found Gigantoraptor to be a Caenagnathid.[11]
| Caenagnathidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology[]
Growth and development[]

A histological analysis performed on the holotype fibula by Xu and colleagues in 2007 showed that the specimen had 7 lines of arrested growth (LAGs, or growth rings) as preserved. As several LAGs were missing, the team used inverse calculations to determine them, resulting in a total of 11. The amount of LAGs indicate the holotype was around 11 years old at the time of death and had an average growth of 128–140 kg (282–309 lb) per year. The performed histology also suggests that this individual was an adult, as there was extensive development of the secondary osteons, tightly packed LAGs close to the periphery of the bone, and a poorly vascularized layer of lamellar bone in the outer circumferential layer, which indicates a relatively sloppy growth rate. However, given that the fourth to seventh sets of LAGs were not very tightly packed, Xu and colleagues inferred that the individual reached its young adulthood at the age of 7. Moreover, the specimen was found to still be a relatively young adult and thereby it would have grown larger when it reached the adult stage. This indicates that Gigantoraptor had an growth rate much faster than most large non-avian theropods, such as tyrannosaurids.[1]
Another unusual development in Gigantoraptor are the elongated hindlimbs. Xu and colleagues pointed out the larger the theropod becomes, the shorter and stockier the legs become in order to withstand the increasing mass. This is contrasted by Gigantoraptor with both femur and tibia being over 1 m (3.3 ft), a combination that is not usually found in theropods of this size. The elongation of these limbs indicates that it was one of the most cursorial—an animal adapted to run or maintain certain speed for a large amount of time—large theropods. Furthermore, Gigantoraptor had an extensively pneumatized vertebral column, which may have aided in weight-loss. The team also suggested that Gigantoraptor might have been "naked" because it is 300 times as heavy as species like Caudipteryx, and large animals tend to rely more on mass for temperature regulation, leading to the loss of insulating coverings found on their smaller relatives. However, they suggested that at least arm feathers were probably still present on Gigantoraptor, since their primary functions, such as display behaviour and covering the eggs while brooding, are not related to the regulation of body heat.[1]
Feeding[]

In 2017, Waisum Ma and colleagues re-examined the preserved jaws of Gigantoraptor and found that it had the deepest beak among caenagnathids and a relatively different diet from other oviraptorosaurs. For instance, the articular region of the jaws of Gigantoraptor show convergence to those of the modern tuatara. The articular region in this reptile enables propalinal jaw movement (a forward and backward movement) during feeding, and based on the similarities between their jaws, Ma and colleagues suggested that Gigantoraptor could have employed propalinal jaw movement as well. This movement aided with the sharp shelves of the dentary may indicate a shearing bite capable of cutting through plants (and potentially meat). This is comparable to other caenagnathids and contrasting with the jaws of oviraptorids, whose jaws seem better suited for crushing food. The depth of the lower jaw indicates the presence of a large tongue, which could have assisted the animal in food proccesing, and possibly improving the feeding efficiency.[3]
As the shape and size of the rostrum in large herbivores is related to dietary habits, the strongly U-shaped dentary of Gigantoraptor may suggest that this oviraptorosaur was a highly non-selective feeder, an adaptation that could have been useful in the variable environmental settings of the Iren Dabasu Formation. The team based this hypothesis on comparisons with several herbivorous animals, including other dinosaurs. Whereas sharply developed and narrow rostra are more correlated with a selective diet, such as the case of some ceratopsians, wide and rounded rostra are more linked with a generalist diet, as seen on the morphology of Gigantoraptor, Euoplocephalus and hadrosaurids. Ma and colleagues also pointed out that its large size, and therefore greater energy requirements, could have influenced its dietary habits as larger animals tend to consume lower quality food since it is available in larger amounts on the surroundings, and has a more stable supply. However, it is not known whether the gut of Gigantoraptor was specialized to process this greater ingestion. The team could not disregard an occasional carnivorous diet as the strong beak together with a propalinal jaw movement may have allowed meat processing.[3]
Reproduction[]

The existence of giant oviraptorosaurians, such as Gigantoraptor, explains several earlier Asian finds of very large, up to 53 centimetres long, oviraptorosaurian eggs, assigned to the oospecies Macroelongatoolithus carlylensis. These were laid in enormous rings with a diameter of three metres. The presence of Macroelongatoolithus in North America indicates that gigantic oviraptorosaurs were present there as well, though no fossil skeletal remains have been found.[12]
In 2018, Kohei Tanaka and team examined the egg clutches of numerous oviraptorosaur specimens, including egg clutches of Macroelongatoolithus, in order to correlate the nest configuration and body size to incubation behaviour. Their results showed that eggshell porosity indicates that the eggs of almost certainly all oviraptorosaurs were exposed in the nest without an external covering. Though most oviraptorosaur nests have eggs arranged in a circular fashion, the morphology of the nest is different in smaller and larger species in that the center of the nest is highly reduced in the former species, and becomes significantly larger in the latter species. This nest configuration suggest that whereas smallest oviraptorosaurs probably sat directly on the eggs, a large, Gigantoraptor-sized animal likely sat on the area devoid of eggs. Tanaka and colleagues pointed out that this adaption was beneficial to avoid egg-crushing and could have allowed some body-contact during incubation in these giant oviraptorosaurs.[13]
Paleoenvironment[]

Gigantoraptor is known from the Iren Dabasu Formation of Inner Mongolia. This formation has been dated to 95.8 ± 6.2 million years ago based on U–Pb and stratigraphic analyses.[14] The environments around this formation were highly humid, existing a large braided river valley with floodplains. A semi-arid climate is also indicated by caliche-based sedimentation. The Iren Dabasu Formation had extensive vegetation and foliage, evidenced in the paleosol development, the numerous herbivorous dinosaurs remains that were found in both the river channel and the floodplain sediments. A rich diversity of charophytes and ostracods inhabited the floodplain systems.[15]
The dinosaurian fauna of this formation includes the fast-running tyrannosauroid Alectrosaurus, the ornithomimosaur Archaeornithomimus, the therizinosaurids Erliansaurus and Neimongosaurus, the sauropod Sonidosaurus, and hadrosauroids Bactrosaurus and Gilmoreosaurus.[16][17] Both Iren Dabasu and Bayan Shireh formations have been correlated on the basis of their similar vertebrate fossil record. This correlation may be further supported by the presence of a giant, Gigantoraptor-sized caenagnathid in the Bayan Shireh Formation. However, given that this specimen is highly fragmentary, its attribution to Gigantoraptor can not be confirmed. The presence of the Bayan Shireh caenagnathid and Gigantoraptor in fluvial-based sedimentation formations like the Iren Dabasu and Bayan Shireh formations suggest a preference over mesic habitats in giant oviraptorosaur species, rather than xeric habitats.[18]
See also[]
- Timeline of oviraptorosaur research
References[]
- ^ Jump up to: a b c d e f g h i j k Xing, X.; Tan, Q.; Wang, J.; Zhao, X.; Tan, L. (2007). "A gigantic bird-like dinosaur from the Late Cretaceous of China". Nature. 447 (7146): 844−847. Bibcode:2007Natur.447..844X. doi:10.1038/nature05849. PMID 17565365. S2CID 6649123. Supplementary Information
- ^ Pickrell, N. (2014). Flying Dinosaurs: How fearsome reptiles became birds. NewSouth Publishing. 256 pp.
- ^ Jump up to: a b c d e Ma, W.; Wang, J.; Pittman, M.; Tan, Q.; Tan., L.; Guo, B.; Xu, X. (2017). "Functional anatomy of a giant toothless mandible from a bird-like dinosaur: Gigantoraptor and the evolution of the oviraptorosaurian jaw". Scientific Reports. 7 (1): 16247. Bibcode:2017NatSR...716247M. doi:10.1038/s41598-017-15709-7. PMC 5701234. PMID 29176627.
- ^ Campione, N. E.; Evans, D. C.; Brown, C. M.; Carrano, M. T. (2014). "Body mass estimation in non‐avian bipeds using a theoretical conversion to quadruped stylopodial proportions". Methods in Ecology and Evolution. 5 (9): 913−923. doi:10.1111/2041-210X.12226. hdl:10088/25281.
- ^ Benson, R. B. J.; Campione, N. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014). "Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage". PLOS Biology. 12 (6): e1001896. doi:10.1371/journal.pbio.1001853. hdl:10088/25994. PMC 4011683. PMID 24802911.
- ^ Molina-Pérez, R.; Larramendi, A. (2016). Récords y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. Barcelona, Spain: Larousse. p. 271. ISBN 9788416641154.
- ^ Persons, W. S.; bruh (2014). "Oviraptorosaur tail forms and functions". Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093.
- ^ Lü, J.; Currie, P. J.; Xu, L.; Zhang, X.; Pu, H.; Jia, S. (2013). "Chicken-sized oviraptorid dinosaurs from central China and their ontogenetic implications". Naturwissenschaften. 100 (2): 165−175. Bibcode:2013NW....100..165L. doi:10.1007/s00114-012-1007-0. PMID 23314810. S2CID 206871470.
- ^ Longrich, N. R.; Currie, P. J.; Zhi-Ming, D. (2010). "A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia". Palaeontology. 53 (5): 945–960. doi:10.1111/j.1475-4983.2010.00968.x.
- ^ Lamanna, M. C.; Sues, H. D.; Schachner, E. R.; Lyson, T. R. (2014). "A New Large-Bodied Oviraptorosaurian Theropod Dinosaur from the Latest Cretaceous of Western North America". PLOS ONE. 10 (4): e0125843. doi:10.1371/journal.pone.0092022. PMC 3960162. PMID 24647078.
- ^ Funston, G.; Currie, P. J. (2016). "A new caenagnathid (Dinosauria: Oviraptorosauria) from the Horseshoe Canyon Formation of Alberta, Canada, and a reevaluation of the relationships of Caenagnathidae". Journal of Vertebrate Paleontology. 36 (4): e1160910. doi:10.1080/02724634.2016.1160910. S2CID 131090028.
- ^ Simon, D. J. (2014). "Giant Dinosaur (Theropod) eggs of the Oogenus Macroelongatoolithus (Elongaroolithidae) from Southeastern Idaho: Taxonimic, Paleobiogeographic, and Reproductive implications" (PDF). Cite journal requires
|journal=(help) - ^ Tanaka, K.; Zelenitsky, D. K.; Lü, J.; DeBuhr, C. L.; Yi, L.; Jia, S.; Ding, F.; Xia, M.; Liu, D.; Shen, C.; Chen, R. (2018). "Incubation behaviours of oviraptorosaur dinosaurs in relation to body size". Biology Letters. 14 (5): 20180135. doi:10.1098/rsbl.2018.0135. PMC 6012691. PMID 29769301.
- ^ Guo, Z. X.; Shi, Y. P.; Yang, Y. T.; Jiang, S. Q.; Li, L. B.; Zhao, Z. G. (2018). "Inversion of the Erlian Basin (NE China) in the early Late Cretaceous: Implications for the collision of the Okhotomorsk Block with East Asia" (PDF). Journal of Asian Earth Sciences. 154: 49−66. doi:10.1016/j.jseaes.2017.12.007.
- ^ Van Itterbeeck, J.; Horne, D. J.; Bultynck, P.; Vandenberghe, N. (2005). "Stratigraphy and palaeoenvironment of the dinosaur-bearing Upper Cretaceous Iren Dabasu Formation, Inner Mongolia, People's Republic of China". Cretaceous Research. 26 (4): 699−725. doi:10.1016/j.cretres.2005.03.004.
- ^ Currie, P. J.; Eberth, D. A. (1993). "Palaeontology, sedimentology and palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People's Republic of China". Cretaceous Research. 14 (2): 127−144. doi:10.1006/cres.1993.1011.
- ^ Xing, H.; He, Y.; Li, L.; Xi, D. (2012). "A review on the study of the stratigraphy, sedimentology, and paleontology of the Iren Dabasu Formation, Inner Mongolia". In Wei, D. (ed.). Proceedings of the Thirteenth Annual Meeting of the Chinese Society of Vertebrate Paleontology (in Chinese). Beijing: China Ocean Press. pp. 1−44.
- ^ Tsuihiji, T.; Watabe, M.; Barsbold, R.; Tsogtbaatar, K. (2015). "A gigantic caenagnathid oviraptorosaurian (Dinosauria: Theropoda) from the Upper Cretaceous of the Gobi Desert, Mongolia". Cretaceous Research. 56: 60−65. doi:10.1016/j.cretres.2015.03.007.
External links[]
 Media related to Gigantoraptor at Wikimedia Commons
Media related to Gigantoraptor at Wikimedia Commons Data related to Gigantoraptor at Wikispecies
Data related to Gigantoraptor at Wikispecies- news@nature.com: "Giant bird-like dinosaur found".
- Wired Science Scientists Discover 3,000-Pound Gigantoraptor Dinosaur in Mongolia
- Fleshed-out restoration of a Gigantoraptor erlianensis. Credit: Julius T. Csotonyi
- Yahoo! News: China finds new species of big, bird-like dinosaur
| hide | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
















- Late Cretaceous dinosaurs of Asia
- Caenagnathids
- Fossil taxa described in 2007
- Taxa named by Xu Xing