Neanderthal
| Neanderthal Temporal range: Middle to Late Pleistocene
| |
|---|---|
| |
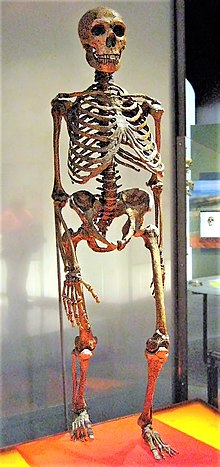
| An approximate reconstruction of a Neanderthal skeleton. The central rib-cage (including the sternum) and parts of the pelvis are from modern humans. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Genus: | Homo |
| Species: | †H. neanderthalensis
|
| Binomial name | |
| †Homo neanderthalensis King, 1864
| |
| |
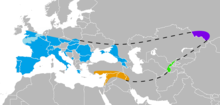
| Known Neanderthal range in Europe (blue), Southwest Asia (orange), Uzbekistan (green), and the Altai Mountains (violet) | |
| Synonyms[6] | |
|
Homo
Palaeoanthropus Protanthropus
Acanthropus
| |
Neanderthals (/niˈændərˌtɑːl, neɪ-, -ˌθɑːl/,[7] also Neandertals, Homo neanderthalensis or Homo sapiens neanderthalensis)[8] are an extinct species or subspecies of archaic humans who lived in Eurasia until about 40,000 years ago.[9][10][11][12] They most likely went extinct due to assimilation into the modern human genome (bred into extinction),[13] great climatic change,[14][15][16] disease,[17][18] or a combination of these factors.[16] They were fully replaced by early European modern humans.
It is unclear when the line of Neanderthals split from that of modern humans; studies have produced various intervals ranging from 315,000[19] to more than 800,000 years ago.[20] The time of divergence of Neanderthals from their ancestor H. heidelbergensis is also unclear. The oldest potential Neanderthal bones date to 430,000 years ago, but the classification remains uncertain.[21] Neanderthals are known from numerous fossils, especially from after 130,000 years ago.[22] The type specimen, Neanderthal 1, was found in 1856 in the Neander Valley in present-day Germany. Researchers depicted Neanderthals as primitive, unintelligent, and brutish for much of the early-20th century. Although knowledge and perception of them has markedly changed since then in the scientific community, the image of the unevolved caveman archetype remains prevalent in popular culture.[23][24]
Neanderthal technology was quite sophisticated. It includes the Mousterian stone-tool industry[25][26] and the ability to create fire[27][28] and build cave hearths,[29][30] make the adhesive birch bark tar,[31] craft at least simple clothes similar to blankets and ponchos,[32] weave,[33] go seafaring through the Mediterranean,[34][35] make use of medicinal plants,[36][37][38] as well as treat severe injuries,[39] store food,[40] and use various cooking techniques such as roasting, boiling,[41] and smoking.[42] Neanderthals made use of a wide array of food, mainly hoofed mammals,[43] but also other megafauna,[23][44] plants,[45][46][47] small mammals, birds, and aquatic and marine resources.[48] Although they were probably apex predators, they still competed with cave bears, cave lions, cave hyaenas, and other large predators.[49] A number of examples of symbolic thought and Palaeolithic art have been inconclusively[50] attributed to Neanderthals, namely possible ornaments made from bird bones[51][52] or shells,[53] collections of unusual objects including crystals and fossils,[54] engravings,[55] music production indicated by the Divje Babe Flute,[56] and Spanish cave-paintings contentiously[57] dated to before 65,000 years ago.[58][59] Some claims of religious beliefs have been made.[60] Neanderthals were likely capable of speech, possibly articulate, although the complexity of their language is not known.[61][62]
Compared with modern humans, Neanderthals had a more robust build and proportionally shorter limbs. Researchers often explain these features as adaptations to conserve heat in a cold climate, but they may also have been adaptations for sprinting in the warmer, forested landscape that Neanderthals often inhabited.[63] Nonetheless, they had cold-specific adaptations, such as specialised body-fat storage[64] and an enlarged nose to warm air[65] (although the nose could have been caused by genetic drift[66]). Average Neanderthal men stood around 165 cm (5 ft 5 in) and women 153 cm (5.02 ft) tall, similar to pre-industrial modern humans.[67] The braincases of Neanderthal men and women averaged about 1,600 cm3 (98 cu in) and 1,300 cm3 (79 cu in) respectively,[68][69][70] which is within the range of the values for modern humans.
The total population of Neanderthals remained low, proliferating weakly harmful gene variants,[71] and precluding effective long-distance networks. Nonetheless, there is evidence of regional cultures and thus of regular communication between communities.[72][73] They may have frequented caves, and moved between caves seasonally.[74] Neanderthals lived in a high-stress environment with high trauma rates, and about 80% died before the age of 40.[75] The 2010 Neanderthal genome project's draft report presented evidence for interbreeding between Neanderthals and modern humans.[76][77][78] It possibly occurred 316–219 thousand years ago,[79] but more likely 100,000 years ago and again 65,000 years ago.[80] Neanderthals also appear to have interbred with Denisovans, a different group of archaic humans, in Siberia.[81][82] Around 1–4% of genomes of Eurasians, Australo-Melanesians, Native Americans, and North Africans is of Neanderthal ancestry, while the inhabitants of sub-Saharan Africa have no detectable Neanderthal ancestry, save possible traces from early Sapiens-to-Neanderthal gene flow and/or more recent back-migration of Eurasians to Africa. In all, about 20% of distinctly Neanderthal gene variants survive today.[83] Although many of the gene variants inherited from Neanderthals may have been detrimental and selected out,[71] Neanderthal introgression appears to have affected the modern human immune system,[84][85][86][87] and is also implicated in several other biological functions and structures,[88] but a large portion appears to be non-coding DNA.[89]
Taxonomy[]
Etymology[]

Neanderthals are named after the valley, the Neandertal, in which the first identified specimen was found. The valley was spelled Neanderthal and the species was spelled Neanderthaler in German until the spelling reform of 1901.[b] The spelling Neandertal for the species is occasionally seen in English, even in scientific publications, but the scientific name, H. neanderthalensis, is always spelled with th according to the principle of priority. The vernacular name of the species in German is always Neandertaler ("inhabitant of the Neander Valley"), whereas Neandertal always refers to the valley.[c] The valley itself was named after the late 17th century German theologian and hymn writer Joachim Neander, who often visited the area.[90] Neanderthal can be pronounced using the /t/ (as in /niˈændərtɑːl/)[93] or the standard English pronunciation of th with the fricative /θ/ (as /niˈændərθɔːl/).[94][95]
Neanderthal 1, the type specimen, was known as the "Neanderthal cranium" or "Neanderthal skull" in anthropological literature, and the individual reconstructed on the basis of the skull was occasionally called "the Neanderthal man".[96] The binomial name Homo neanderthalensis—extending the name "Neanderthal man" from the individual specimen to the entire species, and formally recognising it as distinct from humans—was first proposed by Irish geologist William King in a paper read to the 33rd British Science Association in 1863.[97][98][99] However, in 1864, he recommended that Neanderthals and modern humans be classified in different genera as he compared the Neanderthal braincase to that of a chimpanzee and argued that they were "incapable of moral and [theistic[d]] conceptions".[100]

Research history[]

The first Neanderthal remains—Engis 2 (a skull)—were discovered in 1829 by Dutch naturalist Philippe-Charles Schmerling in the Grottes d'Engis, Belgium, but he thought it was a fossil modern human skull.[101] In 1848, Gibraltar 1 from Forbes' Quarry was presented to the Gibraltar Scientific Society by their Secretary Lieutenant Edmund Henry Réné Flint, but was also thought to be a modern human skull.[102] In 1856, local schoolteacher Johann Carl Fuhlrott recognised bones from Kleine Feldhofer Grotte in Neander Valley—Neanderthal 1 (the holotype specimen)—as distinct from modern humans,[e] and gave them to German anthropologist Hermann Schaaffhausen to study in 1857. It comprised the cranium, thigh bones, right arm, left humerus and ulna, left ilium (hip bone), part of the right shoulder blade, and pieces of the ribs.[100][103] Following Charles Darwin's On the Origin of Species, Fuhlrott and Schaaffhausen argued the bones represented an ancient modern human form;[24][100][104][105] Schaaffhausen, a social Darwinist, believed that humans linearly progressed from savage to civilised, and so concluded that Neanderthals were barbarous cave-dwellers.[24] Fuhlrott and Schaaffhausen met opposition namely from the prolific pathologist Rudolf Virchow who argued against defining new species based on only a single find. In 1872, Virchow erroneously interpreted Neanderthal characteristics as evidence of senility, disease, and malformation instead of archaicness,[106] which stalled Neanderthal research until the end of the century.[24][104]

By the early 20th century, numerous other Neanderthal discoveries were made, establishing H. neanderthalensis as a legitimate species. The most influential specimen was La Chapelle-aux-Saints 1 ("The Old Man") from La Chapelle-aux-Saints, France. French palaeontologist Marcellin Boule authored several publications, among the first to establish palaeontology as a science, detailing the specimen, but reconstructed him as slouching, ape-like, and only remotely related to modern humans. The 1912 'discovery' of Piltdown Man (a hoax), appearing much more similar to modern humans than Neanderthals, was used as evidence that multiple different and unrelated branches of primitive humans existed, and supported Boule's reconstruction of H. neanderthalensis as a far distant relative and an evolutionary dead-end.[24][107][108][109] He fuelled the popular image of Neanderthals as barbarous, slouching, club-wielding primitives; this image was reproduced for several decades and popularised in science fiction works, such as the 1911 The Quest for Fire by J.-H. Rosny aîné and the 1927 The Grisly Folk by H. G. Wells where they are depicted as monsters.[24] In 1911, Scottish anthropologist Arthur Keith reconstructed La Chapelle-aux-Saints 1 as an immediate precursor to modern humans, sitting next to a fire, producing tools, wearing a necklace, and having a more humanlike posture, but this failed to garner much scientific rapport, and Keith later abandoned his thesis in 1915.[24][104][110]

By the middle of the century, based on the exposure of Piltdown Man as a hoax as well as a reexamination of La Chapelle-aux-Saints 1 (who had osteoarthritis which caused slouching in life) and new discoveries, the scientific community began to rework its understanding of Neanderthals. Ideas such as Neanderthal behaviour, intelligence, and culture were being discussed, and a more humanlike image of them emerged. In 1939, American anthropologist Carleton Coon reconstructed a Neanderthal in a modern business suit and hat to emphasise that they would be, more or less, indistinguishable from modern humans had they survived into the present. William Golding's 1955 novel The Inheritors depicts Neanderthals as much more emotional and civilised.[24][23][109] However, Boule's image continued to influence works until the 1960s. In modern-day, Neanderthal reconstructions are often very humanlike.[104][109]
Hybridisation between Neanderthals and early modern humans had been suggested early on,[111] such as by English anthropologist Thomas Huxley in 1890,[112] Danish ethnographer Hans Peder Steensby in 1907,[113] and Coon in 1962.[114] In the early 2000s, supposed hybrid specimens were discovered: Lagar Velho 1[115][116][117][118] and Muierii 1.[119] However, similar anatomy could also have been caused by adapting to a similar environment rather than interbreeding.[89] Neanderthal admixture was found to be present in modern populations in 2010 with the mapping of the first Neanderthal genome sequence.[76] This was based on 3 specimens in Vindija Cave, Croatia, which contained almost 4% archaic DNA (allowing for near complete sequencing of the genome). However, there was approximately 1 error for every 200 letters (base pairs) based on the implausibly high mutation rate, probably due to the preservation of the sample. In 2012, British-American geneticist Graham Coop hypothesised that they instead found evidence of a different archaic human species interbreeding with modern humans, which was disproven in 2013 by the sequencing of a high-quality Neanderthal genome preserved in a toe bone from Denisova Cave, Siberia.[89]
Classification[]
| |||||||||||||||||||||||||||||||||
| 2019 phylogeny based on comparison of ancient proteomes and genomes with those of modern species.[120] |
Neanderthals are hominids in the genus Homo, humans, and generally classified as a distinct species, H. neanderthalensis, although sometimes as a subspecies of modern human as H. sapiens neanderthalensis. This would necessitate the classification of modern humans as H. s. sapiens.[8]
A large part of the controversy stems from the vagueness of the term "species", as it is generally used to distinguish two genetically isolated populations, but admixture between modern humans and Neanderthals is known to have occurred.[8][121] However, the absence of Neanderthal-derived patrilineal Y-chromosome and matrilineal mitochondrial DNA (mtDNA) in modern humans, along with the underrepresentation of Neanderthal X chromosome DNA, could imply reduced fertility or frequent sterility of some hybrid crosses,[78][122][123][124] representing a partial biological reproductive barrier between the groups, and therefore species distinction.[78]
In 2014, geneticist Svante Pääbo described such "taxonomic wars" as unresolvable, "since there is no definition of species perfectly describing the case".[8]
Neanderthals are thought to have been more closely related to Denisovans than to modern humans. Likewise, Neanderthals and Denisovans share a more recent last common ancestor (LCA) than to modern humans, based on nuclear DNA (nDNA). However, Neanderthals and modern humans share a more recent mitochondrial LCA (observable by studying mtDNA). This likely resulted from an interbreeding event subsequent to the Neanderthal/Denisovan split which introduced another mtDNA line. This involved either introgression coming from an unknown archaic human into Denisovans,[81][82][120][125][126] or introgression from an earlier unidentified modern human wave from Africa into Neanderthals.[127]
Evolution[]




It is largely thought that H. heidelbergensis was the last common ancestor of Neanderthals, Denisovans, and modern humans before populations became isolated in Europe, Asia, and Africa, respectively.[129] The taxonomic distinction between H. heidelbergensis and Neanderthals is mostly based on a fossil gap in Europe between 300 and 243 thousand years ago during marine isotope stage 8. "Neanderthals", by convention, are fossils which date to after this gap.[128][23][19] However, 430 thousand year (ka) old bones at Sima de los Huesos could represent early Neanderthals or a closely related group,[21][130][131] and the 400,000 year old Aroeira 3 could represent a transitional phase. Ancestral and derived morphs could have lived concurrently.[132] It is also possible that there was gene flow between Western Europe and Africa during the Middle Pleistocene, obscuring Neanderthal characteristics in such specimens, namely from Ceprano, Italy, and Sićevo Gorge, Serbia.[21] The fossil record is much more complete from 130,000 years ago onwards,[133] and specimens from this period make up the bulk of known Neanderthal skeletons.[134][135] Dental remains from the Italian and sites indicate that Neanderthal dental features had evolved by around 450–430 thousand years ago during the Middle Pleistocene.[136]
There are two main hypotheses regarding the evolution of Neanderthals following the Neanderthal/human split: two-phase and accretion. Two-phase argues a single major environmental event—such as the Saale glaciation—caused European H. heidelbergensis to rapidly increase body size and robustness, as well as undergo a lengthening of the head (phase 1), which then led to other changes in skull anatomy (phase 2).[117] However, Neanderthal anatomy may not have been driven entirely by adapting to cold weather.[63] Accretion holds that Neanderthals slowly evolved over time from the ancestral H. heidelbergensis, divided into 4 stages: early-pre-Neanderthals (MIS 12, Elster glaciation), pre-Neanderthals sensu lato (MIS 11–9, Holstein interglacial), early Neanderthals (MIS 7–5, Saale glaciation–Eemian), and classic Neanderthals sensu stricto (MIS 4–3, Würm glaciation).[128]
Numerous dates for the Neanderthal/human split have been suggested. The date of around 250,000 years ago cites "H. helmei" as being the last common ancestor (LCA), and the split is associated with the Levallois technique of making stone tools. The date of about 400,000 years ago uses H. heidelbergensis as the LCA. 600,000 years ago says that "H. rhodesiensis" was the LCA, which split off into modern human lineage and a Neanderthal/H. heidelbergensis lineage.[137] 800,000 years ago has H. antecessor as the LCA, but different variations of this model would push the date back to 1 million years ago.[21][137] However, a 2020 analysis of H. antecessor enamel proteomes suggests that H. antecessor is related but not a direct ancestor.[138] DNA studies have yielded various results on Neanderthal/human divergence time, such as 538–315,[19] 553–321,[139] 565–503,[140] 654–475,[137] 690–550,[141] 765–550,[21][81] 741–317,[142] and 800–520 thousand years ago;[143] and a dental analysis concluded before 800,000 years ago.[20]
Neanderthals and Denisovans are more closely related to each other than they are to modern humans, meaning the Neanderthal/Denisovan split occurred after their split with modern humans.[21][81][125][144] Assuming a mutation rate of 1x10−9 or 0.5x10−9 per base pair (bp) per year, the Neanderthal/Denisovan split occurred around either 236–190 or 473–381 thousand years ago respectively.[81] Using 1.1x10−8 per generation with a new generation every 29 years, the time is 744,000 years ago. Using 5x10−10 nucleotide sites per year, it is 644,000 years ago. Using the latter dates, the split had likely already occurred by the time hominins spread out across Europe, and unique Neanderthal features had begun evolving by 600–500 thousand years ago.[125] Before splitting, Neanderthal/Denisovans (or "Neandersovans") migrating out of Africa into Europe apparently interbred with an unidentified "superarchaic" human species who were already present there; these superarchaics were the descendants of a very early migration out of Africa around 1.9 mya.[145]
Demographics[]
Range[]

Pre- and early Neanderthals, living before the Eemian interglacial (130,000 years ago), are poorly known and come mostly from Western European sites. From 130,000 years ago onwards, the quality of the fossil record increases dramatically with classic Neanderthals, who are recorded from Western, Central, Eastern, and Mediterranean Europe,[22] as well as Southwest, Central, and Northern Asia up to the Altai Mountains in southern Siberia. Pre- and early Neanderthals, on the other hand, seem to have continuously occupied only France, Spain, and Italy, although some appear to have moved out of this "core-area" to form temporary settlements eastward (although without leaving Europe). Nonetheless, southwestern France has the highest density of sites for pre-, early, and classic Neanderthals.[146]
The southernmost find was recorded at Shuqba Cave, Levant;[147] reports of Neanderthals from the North African Jebel Irhoud[148] and Haua Fteah[149] have been reidentified as H. sapiens. Their easternmost presence is recorded at Denisova Cave, Siberia 85°E; the southeast Chinese Maba Man, a skull, shares several physical attributes with Neanderthals, although these may be the result of convergent evolution rather than Neanderthals extending their range to the Pacific Ocean.[150] The northernmost bound is generally accepted to have been 55°N, with unambiguous sites known between 50–53°N, although this is difficult to assess because glacial advances destroy most human remains, and palaeoanthropologist Trine Kellberg Nielsen has argued that a lack of evidence of Southern Scandinavian occupation is (at least during the Eemian interglacial) due to the former explanation and a lack of research in the area.[151][152] Middle Palaeolithic artefacts have been found up to 60°N on the Russian plains,[153][154][155] but these are more likely attributed to modern humans.[156] A 2017 study claimed the presence of Homo at the 130,000 year old Californian Cerutti Mastodon site in North America,[157] but this is largely considered implausible.[158][159][160]
It is unknown how the rapidly fluctuating climate of the last glacial period (Dansgaard–Oeschger events) impacted Neanderthals, as warming periods would produce more favourable temperatures but encourage forest growth and deter megafauna, whereas frigid periods would produce the opposite.[161] However, Neanderthals may have preferred a forested landscape.[63] Populations may have peaked in cold but not extreme intervals, such as marine isotope stages 8 and 6 (respectively 300 and 191 thousand years ago during the Saale glaciation). It is possible their range expanded and contracted as the ice retreated and grew respectively to avoid permafrost areas, residing in certain refuge zones during glacial maxima.[161] In 2021, Israeli anthropologist Israel Hershkovitz and colleagues suggested the 140 to 120 thousand years old Israeli Nesher Ramla remains, which feature a mix of Neanderthal and more ancient H. erectus traits, represent one such source population which recolonised Europe following a glacial period.[162]

Population[]
Like modern humans, Neanderthals probably descended from a very small population with an effective population—the number of individuals who can bear or father children—of 3,000 to 12,000 approximately. However, Neanderthals maintained this very low population, proliferating weakly harmful genes due to the reduced effectivity of natural selection.[71][163] Various studies, using mtDNA analysis, yield varying effective populations,[161] such as about 1,000 to 5,000;[163] 5,000 to 9,000 remaining constant;[164] or 3,000 to 25,000 steadily increasing until 52,000 years ago before declining until extinction.[73] However, all agree on low population,[161] which may have been less than 1/10th of the contemporary human populations in Western Europe[165] possibly because Neanderthals had much lower fertility rates.[166] Estimates giving a total population in the higher tens of thousands[125] are contested.[163] A consistently low population may be explained in the context of the "Boserupian Trap": a population's carrying capacity is limited by the amount of food it can obtain, which in turn is limited by its technology. Innovation increases with population, but if the population is too low, innovation will not occur very rapidly and the population will remain low. This is consistent with the apparent 150,000 year stagnation in Neanderthal lithic technology.[161]
In a sample of 206 Neanderthals, based on the abundance of young and mature adults in comparison to other age demographics, about 80% of them above the age of 20 died before reaching 40. This high mortality rate was probably due to their high-stress environment.[75] However, it has also been estimated that the age pyramids for Neanderthals and contemporary modern humans were the same.[161] Infant mortality was estimated to have been very high for Neanderthals, about 43% in northern Eurasia.[167]

View references

Anatomy[]
Build[]
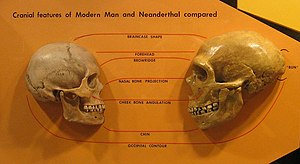
Neanderthals had more robust and stockier builds than modern humans,[67] wider and barrel-shaped rib cages; wider pelvises;[168][23] and proportionally shorter forearms and forelegs.[63][169]
Based on 45 Neanderthal long bones from 14 men and 7 women, the average height was 164 to 168 cm (5 ft 5 in to 5 ft 6 in) for males and 152 to 156 cm (5 ft 0 in to 5 ft 1 in) for females.[67] For comparison, the average height of 28 males and 10 females Upper Palaeolithic humans is respectively 176 cm (5 ft 9 in) and 163 cm (5 ft 4 in), although this decreases by 10 cm (3.9 in) nearer the end of the period based on 21 males and 15 females;[170] and the average in the year 1900 was 163 cm (5 ft 4 in) and 152.7 cm (5.01 ft), respectively.[171] The fossil record shows adult Neanderthals varied from about 147.5 to 177 cm (4 ft 10 in to 5 ft 10 in) in height, although some may have grown much taller.[172] For Neanderthal weight, samples of 26 specimens found an average of 77.6 kg (171 lb) for males and 66.4 kg (146 lb) for females.[173] Using 76 kg (168 lb), the body mass index for Neanderthal males was calculated to be 26.9–28.2, which in modern humans correlates to being overweight. This indicates a very robust build.[67] The Neanderthal LEPR gene concerned with storing fat and body heat production is similar to that of the woolly mammoth, and so was likely an adaptation for cold climate.[64]
The neck vertebrae of Neanderthals are longer and thicker than those of modern humans, lending to stability, possibly due to different head shape and size.[174] Although the Neanderthal thorax (where the ribcage is) was similar in size to modern humans, the longer and straighter ribs would have equated to a widened mid-lower thorax and stronger breathing in the lower thorax, which are indicative of a larger diaphragm and possibly greater lung capacity.[168][175][176] The lung capacity of Kebara 2 was estimated to have been 9.04 L (2.39 US gal). The Neanderthal chest was also more pronounced (expanded front-to-back, or antero-posteriorly). The sacrum (where the pelvis connects to the spine) was more vertically inclined, and was placed lower in relation to the pelvis, causing the spine to be less curved (exhibit less lordosis) and to fold in on itself somewhat (to be invaginated). Such modifications to the spine would have enhanced side-to-side (mediolateral) flexion, better supporting the wider lower thorax. This condition may be normal for Homo, with the condition of a narrower thorax in modern humans being a unique characteristic.[168]
Body proportions are usually cited as being "hyperarctic" as adaptations to the cold, because they are similar to those of human populations which developed in cold climates[177]—the Neanderthal build is most similar to that of Inuit and Siberian Yupiks among modern humans[178]—and shorter limbs equates to higher retention of body heat.[169][177][179] Nonetheless, Neanderthals from more temperate climates—such as Iberia—still retain the "hyperarctic" physique.[180] In 2019, English anthropologist John Stewart and colleagues suggested Neanderthals instead were adapted for sprinting because of evidence of Neanderthals preferring more warmer wooded areas over the colder mammoth steppe, and DNA analysis indicating a higher proportion of fast-twitch muscle fibres in Neanderthals than modern humans. He explained their body proportions and greater muscle mass as adaptations to sprinting as opposed to the endurance-oriented modern human physique,[63] as persistence hunting may only be effective in hot climates where the hunter can run prey to the point of heat exhaustion (hyperthermia). They had longer heel bones,[181] reducing their ability for endurance running, and their shorter limbs would have reduced moment arm at the limbs, allowing for greater rotational force at the wrists and ankles without extra exertion of the rotating muscles at the elbows and knees by increasing the speed at which the muscles contracted, causing faster acceleration.[63] In 1981, American palaeoanthropologist Erik Trinkaus made note of this alternate explanation, but considered it less likely.[169][182]
Face[]

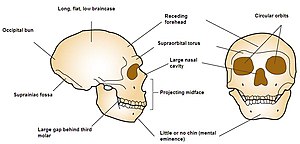
Neanderthals had a reduced chin, sloping forehead, and large nose, which also started somewhat higher on the face than in modern humans. The Neanderthal skull is typically more elongated and less globular than that of modern humans, and features an occipital bun,[184] or "chignon", a protrusion on the back of the skull, although it is within the range of variation for humans who have it. It is caused by the cranial base and temporal bones being placed higher and more towards the front of the skull, and a flatter skullcap.[185] They also had larger eyes likely to adapt to the low-light environment.[186]
The large Neanderthal nose and paranasal sinuses have generally been explained as having warmed air as it entered the lungs and retained moisture ("nasal radiator" hypothesis);[187] but sinuses are generally reduced in cold-adapted creatures, and it may have been that the large nose was caused instead by genetic drift. Also, the sinuses are not grossly large, and are comparable in size to those of modern humans.[66][187] However, sinus size is not an important factor for breathing cold air, and their actual function is unclear, so they may not be a good indicator of evolutionary pressures to evolve such a nose.[188] Further, a computer reconstruction of the Neanderthal nose and predicted soft tissue patterns shows some similarities to those of modern Arctic peoples, potentially meaning the noses of both populations convergently evolved for breathing cold, dry air.[65]
Neanderthals featured a protrusion of the jaw (prognathism), which was once cited as a response to a large bite force evidenced by heavy wearing of Neanderthal front teeth (the "anterior dental loading" hypothesis), but similar wearing trends are seen in contemporary humans. It could also have evolved to fit larger teeth in the jaw, which would better resist wear and abrasion,[187][189] and the increased wear on the front teeth compared to the back teeth probably stems from repetitive use. Neanderthal dental wear patterns are most similar to those of modern Inuit.[187] The incisors are large and shovel-shaped, and, compared to modern humans, there was an unusually high frequency of taurodontism, a condition where the molars are bulkier due to an enlarged pulp (tooth core). Taurodontism was once thought to have been a distinguishing characteristic of Neanderthals which lent some mechanical advantage or stemmed from repetitive use, but was more likely simply a product of genetic drift.[190] The bite force of Neanderthals and modern humans is now thought to be about the same,[187] about 285 N (64 lbf) and 255 N (57 lbf) in modern human males and females, respectively.[191]

Brain[]
The Neanderthal braincase averages 1,600 cm3 (98 in3) for males and 1,300 cm3 (79 in3) for females,[68][69][70] within the possible range of modern humans,[193] which is, on average, 1,270 cm3 (78 in3) for males and 1,130 cm3 (69 in3) for females in present-day.[194] For 28 modern human specimens from 190 to 25 thousand years ago, the average was about 1,478 cc (90.2 cu in) disregarding sex, and modern human brain size is suggested to have decreased since the Upper Palaeolithic.[195] The largest Neanderthal brain, Amud 1, was calculated to be 1,736 cm3 (105.9 in3), one of the largest ever recorded in hominids.[70] Both Neanderthal and human infants measure about 400 cm3 (24 in3).[196]
In Neanderthals, the occipital lobe—operating vision—was much larger than in modern humans, and, similarly, they had larger eyes, probably as an adaptation to lower light conditions in Europe. More brain tissue was devoted to bodily maintenance and control, and, consequently, the cognitive areas of the brain were proportionally smaller than in modern humans,[186] including the cerebellum (operating muscle memory, and possibly language, attention, working memory, social abilities, and thought), the parietal lobes (visuospatial function and episodic memory), the temporal lobes (language comprehension and associations with emotions), the orbitofrontal cortex (decision making), and the olfactory bulb (sense of smell).[197]
Hair and skin colour[]
The lack of sunlight most likely led to the proliferation of lighter skin in Neanderthals,[198] although light skin in modern Europeans was not particularly prolific until perhaps the Bronze Age.[199] Genetically, BNC2 was present in Neanderthals, which is associated with light skin colour; however, a second variation of BNC2 was also present, which is associated with darker skin colour in the UK Biobank.[198] It is likely Neanderthal skin colour varied from region to region. The DNA of three Croatian Neanderthals shows they had darker hair, skin, and eye colour than modern Europeans.[200]
In modern humans, skin and hair colour is regulated by the melanocyte-stimulating hormone—which increases the proportion of eumelanin (black pigment) to phaeomelanin (red pigment)—which is encoded by the MC1R gene. There are 5 known variants in modern humans of the gene which cause loss-of-function and are associated with light skin and hair colour, and another unknown variant in Neanderthals (the R307G variant) which could be associated with pale skin and red hair. The R307G variant was identified in a Neanderthal from Monti Lessini, Italy, and possibly Cueva del Sidrón, Spain.[201] However, as in modern humans, red was probably not a very common hair colour because the variant is not present in many other sequenced Neanderthals.[198]
Metabolism[]
Maximum natural lifespan and the timing of adulthood, menopause, and gestation were most likely very similar to modern humans.[161] However, it has been hypothesised that Neanderthals matured faster than modern humans based on the growth rates of teeth and tooth enamel,[202][203] although this is not backed up by age biomarkers.[75] The main differences in maturation are the atlas bone in the neck as well as the middle thoracic vertebrae fused about 2 years later in Neanderthals than in modern humans, but this was more likely caused by a difference in anatomy rather than growth rate.[204][205]
Generally, models on Neanderthal caloric requirements report significantly higher intakes than those of modern humans because they typically assume Neanderthals had higher basal metabolic rates (BMRs) due to higher muscle mass, faster growth rate, and greater body heat production against the cold;[206][207][208] and higher daily physical activity levels (PALs) due to greater daily travelling distances while foraging.[207][208] However, using a high BMR and PAL, American archaeologist Bryan Hockett estimated that a pregnant Neanderthal would have consumed 5,500 calories per day, which would have necessitated a heavy reliance on big game meat; such a diet would have caused numerous deficiencies or nutrient poisonings, so he concluded that these are unfair assumptions to make.[208]
Neanderthals may have been more active during dimmer light conditions rather than broad daylight because they lived in regions with reduced daytime hours, hunted large game (such predators typically hunt at night to enhance ambush tactics), and had large eyes and visual processing neural centres. Genetically, colour blindness (which may enhance mesopic vision) is typically correlated with northern-latitude populations, and the Neanderthals from Vindija Cave, Croatia, had some substitutions in the Opsin genes which could have influenced colour vision. However, the functional implications of these substitutions are inconclusive.[209] Neanderthal-derived alleles near ASB1 and EXOC6 are associated with being an evening person, narcolepsy, and day-time napping.[198]
Pathology[]
Neanderthals suffered a high rate of traumatic injury, with an estimated 79–94% of specimens showing evidence of healed major trauma, of which 37–52% were severely injured, and 13–19% injured before reaching adulthood.[210] One extreme example is Shanidar 1, who shows signs of an amputation of the right arm likely due to a nonunion after breaking a bone in adolescence, osteomyelitis (a bone infection) on the left clavicle, an abnormal gait, vision problems in the left eye, and possible hearing loss[211] (perhaps swimmer's ear).[212] In 1995, Trinkaus estimated that about 80% succumbed to their injuries and died before reaching 40, and thus theorised that Neanderthals employed a risky hunting strategy ("rodeo rider" hypothesis).[75] However, rates of cranial trauma are not significantly different between Neanderthals and Middle Palaeolithic modern humans (although Neanderthals seem to have had a higher mortality risk),[213] there are few specimens of both Upper Palaeolithic modern humans and Neanderthals who died after the age of 40,[166] and there are overall similar injury patterns between them. In 2012, Trinkaus concluded that Neanderthals instead injured themselves in the same way as contemporary humans, such as by interpersonal violence.[214] A 2016 study looking at 124 Neanderthal specimens argued that high trauma rates were instead caused by animal attacks, and found that about 36% of the sample were victims of bear attacks, 21% big cat attacks, and 17% wolf attacks (totalling 92 positive cases, 74%). There were no cases of hyaena attacks, although hyaenas still nonetheless probably attacked Neanderthals, at least opportunistically.[215] Such intense predation probably stemmed from common confrontations due to competition over food and cave space, and from Neanderthals hunting these carnivores.[215]

Low population caused a low genetic diversity and probably inbreeding, which reduced the population's ability to filter out harmful mutations (inbreeding depression). However, it is unknown how this affected a single Neanderthal's genetic burden and, thus, if this caused a higher rate of birth defects than in modern humans.[216] It is known, however, that the 13 inhabitants of Sidrón Cave collectively exhibited 17 different birth defects likely due to inbreeding or recessive disorders.[217] Likely due to advanced age (60s or 70s), La Chapelle-aux-Saints 1 had signs of Baastrup's disease, affecting the spine, and osteoarthritis.[218] Shanidar 1, who likely died at about 30 or 40, was diagnosed with the most ancient case of diffuse idiopathic skeletal hyperostosis (DISH), a degenerative disease which can restrict movement, which, if correct, would indicate a moderately high incident rate for older Neanderthals.[219]
Neanderthals were likely subject to several infectious diseases and parasites. Modern humans likely transmitted diseases to them; one possible candidate is the stomach bacteria Helicobacter pylori.[220] The modern human papillomavirus variant 16A may descend from Neanderthal introgression.[221] A Neanderthal at Cueva del Sidrón, Spain, shows evidence of a gastrointestinal Enterocytozoon bieneusi infection.[222] The leg bones of the French La Ferrassie 1 feature lesions that are consistent with periostitis—inflammation of the tissue enveloping the bone—likely a result of hypertrophic osteoarthropathy, which is primarily caused by a chest infection or lung cancer.[223] Neanderthals had a lower cavity rate than modern humans, despite some populations consuming typically cavity-causing foods in great quantity, which could indicate a lack of cavity-causing oral bacteria, namely Streptococcus mutans.[224]
Two 250,000-year-old Neanderthal children from Payré, France, present the earliest known cases of lead exposure of any hominin. They were exposed on two distinct occasions either by eating or drinking contaminated food or water, or inhaling lead-laced smoke from a fire. There are two lead mines within 25 km (16 mi) of the site.[225]
Culture[]
Social structure[]
Group dynamics[]

Neanderthals likely lived in more sparsely distributed groups than contemporary modern humans,[161] but group size is thought to have averaged 10 to 30 individuals, similar to modern hunter-gatherers.[29] Reliable evidence of Neanderthal group composition comes from Cueva del Sidrón, Spain, and the footprints at Le Rozel, France:[172] the former shows 7 adults, 3 adolescents, 2 juveniles, and an infant;[226] whereas the latter, based on footprint size, shows a group of 10 to 13 members where juveniles and adolescents made up 90%.[172]
A Neanderthal child's teeth analysed in 2018 showed it was weaned after 2.5 years, similar to modern hunter gatherers, and was born in the spring, which is consistent with modern humans and other mammals whose birth cycles coincide with environmental cycles.[225] Indicated from various ailments resulting from high stress at a low age, such as stunted growth, British archaeologist Paul Pettitt hypothesised that children of both sexes were put to work directly after weaning;[167] and Trinkaus said that, upon reaching adolescence, an individual may have been expected to join in hunting large and dangerous game.[75] However, the bone trauma is comparable to modern Inuit, which could suggest a similar childhood between Neanderthals and contemporary modern humans.[227] Further, such stunting may have also resulted from harsh winters and bouts of low food resources.[225]
Sites showing evidence of no more than three individuals may have represented nuclear families or temporary camping sites for special task groups (such as a hunting party).[29] Bands likely moved between certain caves depending on the season, indicated by remains of seasonal materials such as certain foods, and returned to the same locations generation after generation. Some sites may have been used for over 100 years.[228] Cave bears may have greatly competed with Neanderthals for cave space, and there is a decline in cave bear populations starting 50,000 years ago onwards (although their extinction occurred well after Neanderthals had died out).[229][230] Although Neanderthals are generally considered to have been cave dwellers, with 'home base' being a cave, open-air settlements near contemporaneously inhabited cave systems in the Levant could indicate mobility between cave and open-air bases in this area. Evidence for long-term open-air settlements is known from the 'Ein Qashish site in Israel,[231][232] and Moldova I in Ukraine. Although Neanderthals appear to have had the ability to inhabit a range of environments—including plains and plateaux—open-air Neanderthals sites are generally interpreted as having been used as slaughtering and butchering grounds rather than living spaces.[74]
Inter-group relations[]
Canadian ethnoarchaeologist Brian Hayden calculated a self-sustaining population that avoids inbreeding to consist of about 450–500 individuals, which would necessitate these bands to interact with 8–53 other bands, but more likely the more conservative estimate given low population density.[29] Analysis of the mtDNA of the Neanderthals of Cueva del Sidrón, Spain, showed that the three adult men belonged to the same maternal lineage, while the three adult women belonged to different ones. This suggests a patrilocal residence (that a woman moved out of her group to live with her husband).[233] However, the DNA of a Neanderthal from Denisova Cave, Russia, shows that she had an inbreeding coefficient of 1⁄8 (her parents were either half-siblings with a common mother, double first cousins, an uncle and niece or aunt and nephew, or a grandfather and granddaughter or grandmother and grandson)[81] and the inhabitants of Cueva del Sidrón show several defects, which may have been caused by inbreeding or recessive disorders.[217]
Considering most Neanderthal artefacts were sourced no more than 5 km (3.1 mi) from the main settlement, Hayden considered it unlikely these bands interacted very often,[29] and mapping of the Neanderthal brain and their small group size and population density could indicate that they had a reduced ability for inter-group interaction and trade.[186] However, a few Neanderthal artefacts in a settlement could have originated 20, 30, 100, and 300 km (12.5, 18.5, 60, and 185 mi) away. Based on this, Hayden also speculated that macro-bands formed which functioned much like those of the low-density hunter-gatherer societies of the Western Desert of Australia. Macro-bands collectively encompass 13,000 km2 (5,000 sq mi), with each band claiming 1,200–2,800 km2 (460–1,080 sq mi), maintaining strong alliances for mating networks or to cope with leaner times and enemies.[29] Similarly, British anthropologist Eiluned Pearce and Cypriot archaeologist Theodora Moutsiou speculated that Neanderthals were possibly capable of forming geographically expansive ethnolinguistic tribes encompassing upwards of 800 people, based on the transport of obsidian up to 300 km (190 mi) from the source compared to trends seen in obsidian transfer distance and tribe size in modern hunter-gatherers. However, likely due to a significantly lower population, their model also reported that Neanderthals would not have been as efficient at maintaining long-distance networks as contemporary humans.[234] Hayden noted an apparent cemetery of six or seven individuals at La Ferrassie, France, which, in modern humans, is typically used as evidence of a corporate group which maintained a distinct social identity and controlled some resource, trading, manufacturing, and so on. La Ferrassie is also located in one of the richest animal-migration routes of Pleistocene Europe.[29]

Genetic analysis indicates there were at least 3 distinct geographical groups—Western Europe, the Mediterranean coast, and east of the Caucasus—with some migration among these regions.[73] Post-Eemian Western European Mousterian lithics can also be broadly grouped into 3 distinct macro-regions: Acheulean-tradition Mousterian in the southwest, Micoquien in the northeast, and Mousterian with bifacial tools (MBT) in between the former two. MBT may actually represent the interactions and fusion of the two different cultures.[72] Southern Neanderthals exhibit regional anatomical differences from northern counterparts: a less protrusive jaw, a shorter gap behind the molars, and a vertically higher jawbone.[235] These all instead suggest Neanderthal communities regularly interacted with neighbouring communities within a region, but not as often beyond.[72]
Nonetheless, over long periods of time, there is evidence of large-scale cross-continental migration. Early specimens from Mezmaiskaya Cave in the Caucasus[126] and Denisova Cave in the Siberian Altai Mountains[79] differ genetically from those found in Western Europe, whereas later specimens from these caves both have genetic profiles more similar to Western European Neanderthal specimens than to the earlier specimens from the same locations, suggesting long-range migration and population replacement over time.[79][126] Similarly, artefacts and DNA from and Okladnikov Caves, also in the Altai Mountains, resemble those of eastern European Neanderthal sites about 3,000–4,000 km (1,900–2,500 mi) away more than they do artefacts and DNA of the older Neanderthals from Denisova Cave, suggesting two distinct migration events into Siberia.[236] Neanderthals seem to have suffered a major population decline during MIS 4 (71–57 thousand years ago), and the distribution of the Micoquian tradition could indicate that Central Europe and the Caucasus were repopulated by communities from a refuge zone either in eastern France or Hungary (the fringes of the Micoquian tradition) who dispersed along the rivers Prut and Dniester.[237]
There is also evidence of inter-group conflict: a skeleton from , France, showing a healed fracture on top of the skull apparently caused by a deep blade wound,[238] and another from Shanadir Cave, Iraq, found to have a rib lesion characteristic of projectile weapon injuries.[239]
Social hierarchy[]
It is sometimes suggested, since they were hunters of challenging big game and lived in small groups, there was no sexual division of labour as seen in modern hunter-gatherer societies. That is, men, women, and children all had to be involved in hunting, instead of men hunting with women and children foraging. However, with modern hunter-gatherers, the higher the meat dependency, the higher the division of labour.[29] Further, tooth-wearing patterns in Neanderthal men and women suggest they commonly used their teeth for carrying items, but men exhibit more wearing on the upper teeth, and women the lower, suggesting some cultural differences in tasks.[240]
It is controversially proposed that some Neanderthals wore decorative clothing or jewellery—such as a leopard skin or raptor feathers—to display elevated status in the group. Hayden postulated that the small number of Neanderthal graves found was because only high-ranking members would receive an elaborate burial, as is the case for some modern hunter-gatherers.[29] Trinkaus suggested that elderly Neanderthals were given special burial rites for lasting so long given the high mortality rates.[75] Alternatively, many more Neanderthals may have received burials, but the graves were infiltrated and destroyed by bears.[241] Given that 20 graves of Neanderthals aged under 4 have been found—over a third of all known graves—deceased children may have received greater care during burial than other age demographics.[227]
Looking at Neanderthal skeletons recovered from several natural rock shelters, Trinkaus said that, although Neanderthals were recorded as bearing several trauma-related injuries, none of them had significant trauma to the legs that would debilitate movement. He suggested that self worth in Neanderthal culture derived from contributing food to the group; a debilitating injury would remove this self-worth and result in near-immediate death, and individuals who could not keep up with the group while moving from cave to cave were left behind.[75] However, there are examples of individuals with highly debilitating injuries being nursed for several years, and caring for the most vulnerable within the community dates even further back to H. heidelbergensis.[39][227] Especially given the high trauma rates, it is possible that such an altruistic strategy ensured their survival as a species for so long.[39]
Food[]
Hunting and gathering[]

Neanderthals were once thought of as scavengers, but are now considered to have been apex predators.[242][243] In 1980, it was hypothesised that two piles of mammoth skulls at La Cotte de St Brelade, Jersey, at the base of a gulley were evidence of mammoth drive hunting (causing them to stampede off a ledge),[244] but this is contested.[245] Living in a forested environment, Neanderthals were likely ambush hunters, getting close to and attacking their target—a prime adult—in a short burst of speed, thrusting in a spear at close quarters.[63][246] Younger or wounded animals may have been hunted using traps, projectiles, or pursuit.[246] Nonetheless, they were able to adapt to a variety of habitats.[48][245] They appear to have eaten predominantly what was abundant within their immediate surroundings,[50] with steppe-dwelling communities (generally outside of the Mediterranean) subsisting almost entirely on meat from large game, forest-dwelling communities consuming a wide array of plants and smaller animals, and waterside communities gathering aquatic resources. Contemporary humans, in contrast, seem to have used more complex food extraction strategies and generally had a more diverse diet.[247] Nonetheless, Neanderthals still would have had to have eaten a varied enough diet to prevent nutrient deficiencies and protein poisoning, especially in the winter when they presumably ate mostly lean meat. Any food with high contents of other essential nutrients not provided by lean meat would have been vital components of their diet, such as fat-rich brains,[39] carbohydrate-rich and abundant underground storage organs (including roots and tubers),[248] or, like modern Inuit, the stomach contents of herbivorous prey items.[249]
For meat, they appear to have fed predominantly on hoofed mammals, namely red deer and reindeer as these two were the most abundant game,[43] but also on other Pleistocene megafauna such as ibex, wild boar, aurochs, mammoth, straight-tusked elephant, woolly rhinoceros, and so on.[23][44][250] There is evidence of directed cave and brown bear hunting both in and out of hibernation, as well as butchering.[251] Analysis of Neanderthal bone collagen from Vindija Cave, Croatia, shows nearly all of their protein needs derived from animal meat.[44] Some caves show evidence of regular rabbit and tortoise consumption. At Gibraltar sites, there are remains of 143 different bird species, many ground-dwelling such as the common quail, corn crake, woodlark, and crested lark.[48] Neanderthals also exploited marine resources on the Iberian, Italian, and Peloponnesian Peninsulas, where they waded or dived for shellfish,[48][252][253] as early as 150,000 years ago at , Spain, similar to the fishing record of modern humans.[254] At Vanguard Cave, Gibraltar, the inhabitants consumed Mediterranean monk seal, short-beaked common dolphin, common bottlenose dolphin, Atlantic bluefin tuna, sea bream, and purple sea urchin;[48][255] and at Gruta da Figueira Brava, Portugal, there is evidence of large-scale harvest of shellfish, crabs, and fish.[256] Evidence of freshwater fishing was found in Grotte di Castelcivita, Italy, for trout, chub, and eel;[253] , France, for chub and European perch; Payré, France;[257] and , Russia, for Black Sea salmon.[258]
Edible plant and mushroom remains are recorded from several caves.[46] Neanderthals from Cueva del Sidrón, Spain, based on dental tartar, likely had a meatless diet of mushrooms, pine nuts, and moss, indicating they were forest foragers.[222] Remnants from Amud Cave, Israel, indicates a diet of figs, palm tree fruits, and various cereals and edible grasses.[47] Several bone traumas in the leg joints could possibly suggest habitual squatting, which, if the case, was likely done while gathering food.[259] Dental tartar from Grotte de Spy, Belgium, indicates the inhabitants had a meat-heavy diet including woolly rhinoceros and mouflon sheep, while also regularly consuming mushrooms.[222] Neanderthal faecal matter from El Salt, Spain, dated to 50,000 years ago—the oldest human faecal matter remains recorded—show a diet mainly of meat but with a significant component of plants.[260] Evidence of cooked plant foods—mainly legumes and, to a far lesser extent, acorns—was discovered in Kebara Cave, Israel, with its inhabitants possibly gathering plants in spring and fall and hunting in all seasons except fall, although the cave was probably abandoned in late summer to early fall.[37] At Shanidar Cave, Iraq, Neanderthals collected plants with various harvest seasons, indicating they scheduled returns to the area to harvest certain plants, and that they had complex food-gathering behaviours for both meat and plants.[45]
Food preparation[]
Neanderthals probably could employ a wide range of cooking techniques, such as roasting, and they may have been able to heat up or boil soup, stew, or animal stock.[41] The abundance of animal bone fragments at settlements may indicate the making of fat stocks from boiling bone marrow, possibly taken from animals that had already died of starvation. These methods would have substantially increased fat consumption, which was a major nutritional requirement of communities with low carbohydrate and high protein intake.[41][261] Neanderthal tooth size had a decreasing trend after 100,000 years ago, which could indicate an increased dependence on cooking or the advent of boiling, a technique that would have softened food.[262]

At Cueva del Sidrón, Spain, Neanderthals likely cooked and possibly smoked food,[42] as well as used certain plants—such as yarrow and camomile—as flavouring,[41] although these plants may have instead been used for their medicinal properties.[36] At Gorham's Cave, Gibraltar, Neanderthals may have been roasting pinecones to access pine nuts.[48]
At Grotte du Lazaret, France, a total of 23 red deer, 6 ibexes, 3 aurochs, and 1 roe deer appear to have been hunted in a single autumn hunting season, when strong male and female deer herds would group together for rut. The entire carcasses seem to have been transported to the cave and then butchered. Because this is such a large amount of food to consume before spoilage, it is possible these Neanderthals were curing and preserving it before winter set in. At 160,000 years old, it is the oldest potential evidence of food storage.[40] The great quantities of meat and fat which could have been gathered in general from typical prey items (namely mammoths) could also indicate food storage capability.[263] With shellfish, Neanderthals needed to eat, cook, or in some manner preserve them soon after collection, as shellfish spoils very quickly. At Cueva de los Aviones, Spain, the remains of edible, algae eating shellfish associated with the alga Jania rubens could indicate that, like some modern hunter gatherer societies, harvested shellfish were held in water-soaked algae to keep them alive and fresh until consumption.[264]
Competition[]

Competition from large Ice Age predators was rather high. Cave lions likely targeted horses, large deer and wild cattle; and leopards primarily reindeer and roe deer; which heavily overlapped with Neanderthal diet. To defend a kill against such ferocious predators, Neanderthals may have engaged in a group display of yelling, arm waving, or stone throwing; or quickly gathered meat and abandoned the kill. However, at Grotte de Spy, Belgium, the remains of wolves, cave lions, and cave bears—which were all major predators of the time—indicate Neanderthals hunted their competitors to some extent.[49]
Neanderthals and cave hyaenas may have exemplified niche differentiation, and actively avoided competing with each other. Although they both mainly targeted the same groups of creatures—deer, horses, and cattle—Neanderthals mainly hunted the former and cave hyaenas the latter two. Further, animal remains from Neanderthal caves indicate they preferred to hunt prime individuals, whereas cave hyaenas hunted weaker or younger prey, and cave hyaena caves have a higher abundance of carnivore remains.[43] Nonetheless, there is evidence that cave hyaenas stole food and leftovers from Neanderthal campsites and scavenged on dead Neanderthal bodies.[265]
Cannibalism[]
There are several instances of Neanderthals practising cannibalism across their range.[266][267] The first example came from Krapina, Croatia, in 1899,[109] and other examples were found at Cueva del Sidrón[235] and Zafarraya in Spain; and the French ,[268] , and La Quina. For the five cannibalised Neanderthals at the Grottes de Goyet, Belgium, there is evidence that the upper limbs were disarticulated, the lower limbs defleshed and also smashed (likely to extract bone marrow), the chest cavity disemboweled, and the jaw dismembered. There is also evidence that the butchers used some bones to retouch their tools. The processing of Neanderthal meat at Grottes de Goyet is similar to how they processed horse and reindeer.[266][267] About 35% of the Neanderthals at Marillac-le-Franc, France, show clear signs of butchery, and the presence of digested teeth indicates that the bodies were abandoned and eaten by scavengers, likely hyaenas.[269]
These cannibalistic tendencies have been explained as either ritual defleshing, pre-burial defleshing (to prevent scavengers or foul smell), an act of war, or simply for food. Due to a small number of cases, and the higher number of cut marks seen on cannibalised individuals than animals (indicating inexperience), cannibalism was probably not a very common practice, and it may have only been done in times of extreme food shortages as in some cases in recorded human history.[267]
Art[]
Personal adornment[]
Neanderthal are known to have used ochre, a clay earth pigment. Ochre is well-documented from 60 to 45 thousand years ago in Neanderthal sites, with the earliest example dating to 250–200 thousand years ago from , the Netherlands (a similar timespan to the ochre record of H. sapiens).[270] It has been hypothesised to have functioned as body paint, and analyses of pigments from Pech de l’Azé, France, indicates they were applied to soft materials (such as a hide or human skin).[271] However, modern hunter gatherers, in addition to body paint, also use ochre for medicine, for tanning hides, as a food preservative, and as an insect repellent, so its use as decorative paint for Neanderthals is speculative.[270] Containers apparently used for mixing ochre pigments were found in , Romania, which could indicate modification of ochre for solely aesthetic purposes.[272]

Neanderthals are known to have collected uniquely shaped objects and are suggested to have modified them into pendants, such as a fossil Aspa marginata sea snail shell possibly painted red from , Italy, transported over 100 km (62 mi) to the site about 47,500 years ago;[273] 3 shells, dated to about 120–115 thousand years ago, perforated through the umbo belonging to a rough cockle, a Glycymeris insubrica, and a Spondylus gaederopus from Cueva de los Aviones, Spain, the former two associated with red and yellow pigments, and the latter a red-to-black mix of hematite and pyrite; and a king scallop shell with traces of an orange mix of goethite and hematite from Cueva Antón, Spain. The discoverers of the latter two claim that pigment was applied to the exterior to make it match the naturally vibrant inside colouration.[53][264] Excavated from 1949 to 1963 from the French Grotte du Renne, Châtelperronian beads made from animal teeth, shells, and ivory were found associated with Neanderthal bones, but the dating is uncertain and Châtelperronian artefacts may actually have been crafted by modern humans and simply redeposited with Neanderthal remains.[274][275][276][277]

Most notably by Gibraltarian palaeoanthropologists Clive and Geraldine Finlayson, Neanderthals are suggested to have used various bird parts as artistic mediums, specifically black feathers.[278] In 2012, the Finlaysons and colleagues examined 1,699 sites across Eurasia, and argued that raptors and corvids, species not typically consumed by any human species, were overrepresented and show processing of only the wing bones instead of the fleshier torso, and thus are evidence of feather plucking of specifically the large flight feathers for use as personal adornment. They specifically noted the cinereous vulture, red-billed chough, kestrel, lesser kestrel, alpine chough, rook, jackdaw, and the white tailed eagle in Middle Palaeolithic sites.[279] Other birds claimed to present evidence of modifications by Neanderthals are the golden eagle, rock pigeon, common raven, and the bearded vulture.[280] The earliest claim of bird bone jewellery is a number of 130,000 year old white tailed eagle talons found in a cache near Krapina, Croatia, speculated, in 2015, to have been a necklace.[281][282] A similar 39,000-year-old Spanish imperial eagle talon necklace was reported in 2019 at Cova Foradà in Spain, though from the contentious Châtelperronian layer.[283] In 2017, 17 incision-decorated raven bones from the rock shelter, Ukraine, dated to 43–38 thousand years ago were reported. Because the notches are more-or-less equidistant to each other, they are the first modified bird bones that cannot be explained by simple butchery, and for which the argument of design intent is based on direct evidence.[51]
Discovered in 1975, the so-called Mask of la Roche-Cotard, a mostly flat piece of flint with a bone pushed through a hole on the midsection—dated to 32, 40, or 75 thousand years ago[284]—has been purported to resemble the upper half of a face, with the bone representing eyes.[285][286] It is contested whether it represents a face, or if it even counts as art.[287] In 1988, American archaeologist Alexander Marshack speculated that a Neanderthal at , France, wore a leopard pelt as personal adornment to indicate elevated status in the group based on a recovered leopard skull, phalanges, and tail vertebrae.[29][288]
Abstraction[]

As of 2014, 63 purported engravings have been reported from 27 different European and Middle Eastern Lower-to-Middle Palaeolithic sites, of which 20 are on flint cortexes from 11 sites, 7 are on slabs from 7 sites, and 36 are on pebbles from 13 sites. It is debated whether or not these were made with symbolic intent.[55] In 2012, deep scratches on the floor of Gorham's Cave, Gibraltar, were discovered, dated to older than 39,000 years ago, which the discoverers have interpreted as Neanderthal abstract art.[289][290] The scratches could have also been produced by a bear.[241] In 2021, an Irish elk phalanx with five engraved offset chevrons stacked above each other was discovered at the entrance to the Einhornhöhle cave in Germany, dating to about 51,000 years ago.[291]
In 2018, some red-painted dots, disks, lines, and hand stencils on the cave walls of the Spanish La Pasiega, Maltravieso, and were dated to be older than 66,000 years ago, at least 20,000 years prior to the arrival of modern humans in Western Europe. This would indicate Neanderthal authorship, and similar iconography recorded in other Western European sites—such as Les Merveilles, France, and Cueva del Castillo, Spain—could potentially also have Neanderthal origins.[58][59][292] However, the dating of these Spanish caves, and thus attribution to Neanderthals, is contested.[57]
Neanderthals are known to have collected a variety of unusual objects—such as crystals or fossils—without any real functional purpose or any indication of damage caused by use. It is unclear if these objects were simply picked up for their aesthetic qualities, or if some symbolic significance was applied to them. These items are mainly quartz crystals, but also other minerals such as cerussite, iron pyrite, calcite, and galena. A few findings feature modifications, such as a mammoth tooth with an incision and a fossil nummulite shell with a cross etched in from Tata, Hungary; a large slab with 18 cupstones hollowed out from a grave in La Ferrassie, France;[54] and a geode from Peștera Cioarei, Romania, coated with red ochre.[293] A number of fossil shells are also known from French Neanderthals sites, such as a rhynchonellid and a from Combe Grenal; a belemnite beak from ; a polyp from ; a sea urchin from La Gonterie-Boulouneix; and a rhynchonella, feather star, and belemnite beak from the contentious Châtelperronian layer of Grotte du Renne.[54]
Music[]

Purported Neanderthal bone flute fragments made of bear long bones were reported from Potočka zijalka, Slovenia, in the 1920s, and Istállós-kői-barlang, Hungary,[294] and , Slovenia, in 1985; but these are now attributed to modern human activities.[295][296] The 1995 43 thousand year old Divje Babe Flute from Slovenia has been attributed by some researchers to Neanderthals, and Canadian musicologist Robert Fink said the original flute had either a diatonic or pentatonic musical scale.[297] However, the date also overlaps with modern human immigration into Europe, which means it is also possible it was not manufactured by Neanderthals.[298] In 2015, zoologist Cajus Diedrich argued that it was not a flute at all, and the holes were made by a scavenging hyaena as there is a lack of cut marks stemming from whittling,[296] but in 2018, Slovenian archaeologist Matija Turk and colleagues countered that it is highly unlikely the punctures were made by teeth, and cut marks are not always present on bone flutes.[56]
Technology[]
Despite the apparent 150 thousand year stagnation in Neanderthal lithic innovation,[161] there is evidence that Neanderthal technology was more sophisticated than was previously thought.[61] However, the high frequency of potentially debilitating injuries could have prevented very complex technologies from emerging, as a major injury would have impeded an expert's ability to effectively teach a novice.[210]
Stone tools[]


Neanderthals made stone tools, and are associated with the Mousterian industry.[25] The Mousterian is also associated with North African H. sapiens as early as 315,000 years ago[299] and was found in Northern China about 47–37 thousand years ago.[300] It evolved around 300,000 years ago with the Levallois technique which developed directly from the preceding Acheulean industry (invented by H. erectus about 1.8 mya). Levallois made it easier to control flake shape and size, and as a difficult-to-learn and unintuitive process, the Levallois technique may have been directly taught generation to generation rather than via purely observational learning.[26]
There are distinct regional variants of the Mousterian industry, such as: the Quina and La Ferrassie subtypes of the Charentian industry in southwestern France, Acheulean-tradition Mousterian subtypes A and B along the Atlantic and northwestern European coasts,[301] the Micoquien industry of Central and Eastern Europe and the related Sibiryachikha variant in the Siberian Altai Mountains,[236] the Denticulate Mousterian industry in Western Europe, the racloir industry around the Zagros Mountains, and the flake cleaver industry of Cantabria, Spain, and both sides of the Pyrenees. In the mid-20th century, French archaeologist François Bordes debated against American archaeologist Lewis Binford to explain this diversity (the "Bordes–Binford debate"), with Bordes arguing that these represent unique ethnic traditions and Binford that they were caused by varying environments (essentially, form vs. function).[301] The latter sentiment would indicate a lower degree of inventiveness compared to modern humans, adapting the same tools to different environments rather than creating new technologies.[50] A continuous sequence of occupation is well-documented in Grotte du Renne, France, where the lithic tradition can be divided into the Levallois–Charentian, Discoid–Denticulate (43.3±0.929–40.9±0.719 thousand years ago), Levallois Mousterian (40.2±1.5–38.4±1.3 thousand years ago), and Châtelperronian (40.93±0.393–33.67±0.450 thousand years ago).[302]

There is some debate if Neanderthals had long-ranged weapons.[303][304] A wound on the neck of an African wild ass from , Syria, was likely inflicted by a heavy Levallois-point javelin,[305] and bone trauma consistent with habitual throwing has been reported in Neanderthals.[303][304] Some spear tips from Abri du Maras, France, may have been too fragile to have been used as thrusting spears, possibly suggesting their use as darts.[257]
Organic tools[]
The Châtelperronian in central France and northern Spain is a distinct industry from the Mousterian, and is controversially hypothesised to represent a culture of Neanderthals borrowing (or by process of acculturation) tool-making techniques from immigrating modern humans, crafting bone tools and ornaments. In this frame, the makers would have been a transitional culture between the Neanderthal Mousterian and the modern human Aurignacian.[306][307][308][309][310] The opposing viewpoint is that the Châtelperronian was manufactured by modern humans instead.[311] Abrupt transitions similar to the Mousterian/Châtelperronian could also simply represent natural innovation, like the Quina– transition 50,000 years ago featuring technologies generally associated with modern humans such as bladelets and microliths. Other ambiguous transitional cultures include the Italian Uluzzian industry,[312] and the Balkan Szeletian industry.[313]
Before immigration, the only evidence of Neanderthal bone tools are animal rib lissoirs — which are rubbed against hide to make it more supple or waterproof — although this could also be evidence for modern humans immigrating earlier than expected. In 2013, two 51.4–41.1 thousand year old deer rib lissoirs were reported from Pech-de-l’Azé and the nearby in France.[308][89][89] In 2020, 5 more lissoirs made of aurochs or bison ribs were reported from Abri Peyrony, with one dating to about 51,400 years ago and the other four to 47.7–41.1 thousand years ago. This indicates the technology was in use in this region for a long time. Since reindeer remains were the most abundant, the use of less abundant bovine ribs may indicate a specific preference for bovine ribs. Potential lissoirs have also been reported from Grosse Grotte, Germany (made of mammoth), and Grottes des Canalettes, France (red deer).[314]

The Neanderthals in 10 coastal sites in Italy (namely Grotta del Cavallo and ) and , Greece, are known to have crafted scrapers using smooth clam shells, and possibly hafted them to a wooden handle. They probably chose this clam species because it has the most durable shell. At Grotta dei Moscerini, about 24% of the shells were gathered alive from the seafloor, meaning these Neanderthals had to wade or dive into shallow waters to collect them. At , Italy, in the Campanian volcanic arc, Neanderthals collected the porous volcanic pumice, which, for contemporary humans, was probably used for polishing points and needles. The pumices are associated with shell tools.[253]
At Abri du Maras, France, twisted fibres and a 3-ply inner-bark-fibre cord fragment associated with Neanderthals show that they produced string and cordage, but it is unclear how widespread this technology was because the materials used to make them (such as animal hair, hide, sinew, or plant fibres) are biodegradable and preserve very poorly. This technology could indicate at least a basic knowledge of weaving and knotting, which would have made possible the production of nets, containers, packaging, baskets, carrying devices, ties, straps, harnesses, clothes, shoes, beds, bedding, mats, flooring, roofing, walls, and snares, and would have been important in hafting, fishing, and seafaring. Dating to 52–41 thousand years ago, the cord fragment is the oldest direct evidence of fibre technology, although 115,000-year-old perforated shell beads from Cueva Antón possibly strung together to make a necklace are the oldest indirect evidence.[33][257] In 2020, British archaeologist Rebecca Wragg Sykes expressed cautious support for the genuineness of the find, but pointed out that the string would have been so weak that it would have had limited functions. One possibility is as a thread for attaching or stringing small objects.[315]
The archaeological record shows that Neanderthals commonly used animal hide and birch bark, may have used them to make cooking containers, although this is based largely on circumstantial evidence, because neither fossilize well.[262] It is possible the Neanderthals at Kebara Cave, Israel, used the shells of the spur-thighed tortoise as containers.[316]
At the Italian site, there is evidence they used fire to process boxwood branches to make digging sticks, a common implement in hunter-gatherer societies.[317]
Fire and construction[]
Many Mousterian sites have evidence of fire, some for extended periods of time, though it is unclear if they were capable of starting fire or simply scavenged from naturally occurring wildfires. Indirect evidence of fire starting ability includes pyrite residue on a couple dozen bifaces from late Mousterian (c. 50,000 years ago) northwestern France (which could indicate they were used as percussion fire starters), and collection of manganese dioxide by late Neanderthals which can lower the combustion temperature of wood.[27][28][318] They were also capable of zoning areas for specific activities, such as for knapping, butchering, hearths, and wood storage. Many Neanderthal sites lack evidence for such activity perhaps due to natural degradation of the area over tens of thousands of years, such as by bear infiltration after abandonment of the settlement.[241]
In a number of caves, evidence of hearths has been detected. Neanderthals likely considered air circulation when making hearths as a lack of proper ventilation for a single hearth can render a cave uninhabitable in several minutes.[30] Abric Romaní rock shelter, Spain, indicates eight evenly spaced hearths lined up against the rock wall, likely used to stay warm while sleeping, with one person sleeping on either side of the fire.[29][30] At Cueva de Bolomor, Spain, with hearths lined up against the wall, the smoke flowed upwards to the ceiling, and led to outside the cave. In Grotte du Lazaret, France, smoke was probably naturally ventilated during the winter as the interior cave temperature was greater than the outside temperature; likewise, the cave was likely only inhabited in the winter.[30]

In 1990, two 176,000 year old ring structures made of broken stalagmite pieces, several metres wide, were discovered in a large chamber more than 300 m (980 ft) from the entrance within Grotte de Bruniquel, France. One ring was 6.7 m × 4.5 m (22 ft × 15 ft) with stalagmite pieces averaging 34.4 cm (13.5 in) in length, and the other 2.2 m × 2.1 m (7.2 ft × 6.9 ft) with pieces averaging 29.5 cm (11.6 in). There were also 4 other piles of stalagmite pieces for a total of 112 m (367 ft) or 2.2 t (2.4 short tons) worth of stalagmite pieces. Evidence of the use of fire and burnt bones also suggest human activity. A team of Neanderthals was likely necessary to construct the structure, but the chamber's actual purpose is uncertain. Building complex structures so deep in a cave is unprecedented in the archaeological record, and indicates sophisticated lighting and construction technology, and great familiarity with subterranean environments.[319]
The 44,000 year old Moldova I open-air site, Ukraine, shows evidence of a 7 m × 10 m (23 ft × 33 ft) ring-shaped dwelling made out of mammoth bones meant for long-term habitation by several Neanderthals, which would have taken a long time to build. It appears to have contained hearths, cooking areas, and a flint workshop, and there are traces of woodworking. Upper Palaeolithic modern humans in the Russian plains are thought to have also made housing structures out of mammoth bones.[74]
Bark tar[]
Neanderthal produced the adhesive birch bark tar, perhaps using plant-based resins for hafting.[320] It was long believed that birch bark tar required a complex recipe to be followed, and that it thus showed complex cognitive skills and cultural transmission. However, a 2019 study showed it can be made simply by burning birch bark on smooth vertical surfaces, such as a flat, inclined rock.[31]
Clothes[]
Neanderthals were likely able to survive in a similar range of temperatures as modern humans while sleeping: about 32 °C (90 °F) while naked in the open and windspeed 5.4 km/h (3.4 mph), or 27–28 °C (81–82 °F) while naked in an enclosed space. Since ambient temperatures were markedly lower than this—averaging during the Eemian interglacial 17.4 °C (63.3 °F) in July and 1 °C (34 °F) in January and dropping to as a low as −30 °C (−22 °F) on the coldest days—Danish physicist Bent Sørensen hypothesised that Neanderthals required tailored clothing capable of preventing airflow to the skin. Especially during extended periods of travelling (such as a hunting trip), tailored footwear completely enwrapping the feet may have been necessary.[321]

Nonetheless, as opposed to the bone sewing-needles and stitching awls assumed to have been in use by contemporary modern humans, the only known Neanderthal tools that could have been used to fashion clothes are hide scrapers, which could have made items similar to blankets or ponchos, and there is no direct evidence they could produce fitted clothes.[32][322] Indirect evidence of tailoring by Neanderthals includes the ability to manufacture string, which could indicate weaving ability,[257] and a naturally-pointed horse metatarsal bone from Cueva de los Aviones, Spain, which was speculated to have been used as an awl, perforating dyed hides, based on the presence of orange pigments.[264] Whatever the case, Neanderthals would have needed to cover up most of their body, and contemporary humans would have covered 80–90%.[322][323]
Since human/Neanderthal admixture is known to have occurred in the Middle East, and no modern body louse species descends from their Neanderthal counterparts (body lice only inhabit clothed individuals), it is possible Neanderthals (and/or humans) in hotter climates did not wear clothes, or Neanderthal lice were highly specialised.[323]
Seafaring[]
Remains of Middle Palaeolithic stone tools on Greek islands indicate early seafaring by Neanderthals in the Ionian Sea possibly starting as far back as 200–150 thousand years ago. The oldest stone artefacts from Crete date to 130–107 thousand years ago, Cephalonia 125 thousand years ago, and Zakynthos 110–35 thousand years ago. The makers of these artefacts likely employed simple reed boats and made one-day crossings back and forth.[34] Other Mediterranean islands with such remains include Sardinia, Melos, Alonnisos,[35] and Naxos (although Naxos may have been connected to land),[324] and it is possible they crossed the Strait of Gibraltar.[35] If this interpretation is correct, Neanderthals' ability to engineer boats and navigate through open waters would speak to their advanced cognitive and technical skills.[35][324]
Medicine[]
Given their dangerous hunting and extensive skeletal evidence of healing, Neanderthals appear to have lived lives of frequent traumatic injury and recovery. Well-healed fractures on many bones indicate the setting of splints. Individuals with severe head and rib traumas (which would have caused massive blood loss) indicate they had some manner of dressing major wounds, such as bandages made from animal skin. By-and-large, they appear to have avoided severe infections, indicating good long-term treatment of such wounds.[39]
Their knowledge of medicinal plants was comparable to that of contemporary humans.[39] An individual at Cueva del Sidrón, Spain, seems to have been medicating a dental abscess using poplar—which contains salicylic acid, the active ingredient in aspirin—and there were also traces of the antibiotic-producing Penicillium chrysogenum.[222] They may have also used yarrow and camomile, and their bitter taste—which should act as a deterrent as it could indicate poison—means it was likely a deliberate act.[36] In Kebara Cave, Israel, plant remains which have historically been used for their medicinal properties were found, including the common grape vine, the pistachios of the Persian turpentine tree, ervil seeds, and oak acorns.[37]
Language[]

The degree of language complexity is difficult to establish, but given that Neanderthals achieved some technical and cultural complexity, and interbred with humans, it is reasonable to assume they were at least fairly articulate, comparable to modern humans. A somewhat complex language—possibly using syntax—was likely necessary to survive in their harsh environment, with Neanderthals needing to communicate about topics such as locations, hunting and gathering, and tool-making techniques.[61][325][326] The FOXP2 gene in modern humans is associated with speech and language development. FOXP2 was present in Neanderthals,[327] but not the gene's modern human variant.[328] Neurologically, Neanderthals had an expanded Broca's area—operating the formulation of sentences, and speech comprehension, but out of a group 48 genes believed to affect the neural substrate of language, 11 had different methylation patterns between Neanderthals and modern humans. This could indicate a stronger ability in modern humans than in Neanderthals to express language.[329]
In 1971, cognitive scientist Philip Lieberman attempted to reconstruct the Neanderthal vocal tract and concluded that it was similar to that of a newborn and incapable of producing nasal sounds, due to the large size of the mouth and thus no need for a descended larynx to fit the entire tongue inside the mouth. He claimed that they were anatomically unable to produce the sounds /a/, /i/, /u/, /ɔ/, /g/, and /k/ and thus lacked the capacity for articulate speech, though were still able to speak at a level higher than non-human primates.[330][331][332] However, the lack of a descended larynx does not necessarily equate to a reduced vowel capacity.[333] The 1983 discovery of a Neanderthal hyoid bone—used in speech production in humans—in Kebara 2 which is almost identical to that of humans suggests Neanderthals were capable of speech, and the ancestral Sima de los Huesos hominins had humanlike hyoid and ear bones, which could suggest the early evolution of the modern human vocal apparatus. However, the hyoid does not definitively provide insight into vocal tract anatomy.[62] Subsequent studies reconstruct the Neanderthal vocal apparatus as comparable to that of modern humans, with a similar vocal repertoire.[334] In 2015, Lieberman hypothesized that Neanderthals were capable of syntactical language, although nonetheless incapable of mastering any human dialect.[335]
It is debated if behavioural modernity is a recent and uniquely modern human innovation, or if Neanderthals also possessed it.[336][326][337][52]
Religion[]
Funerals[]


Claims that Neanderthals held funerals for their dead with symbolic meaning[338] are heavily contested and speculative.[339][340][341] Although Neanderthals did bury their dead, at least occasionally—which may explain the abundance of fossil remains—[50] it is not indicative of a religious belief of life after death, as such burial could have also had non-symbolic motivations, such as great emotion[342] or to prevent scavenging.[343]
The debate on Neanderthal funerals has been active since the 1908 discovery of La Chapelle-aux-Saints 1 in a small, artificial hole in a cave in southwestern France, very controversially postulated to have been buried in a symbolic fashion.[339][344][345] Another grave at Shanidar Cave, Iraq, was associated with the pollen of several flowers that may have been in bloom at the time of deposition—yarrow, centaury, ragwort, grape hyacinth, joint pine, and hollyhock.[346] The medicinal properties of the plants led American archaeologist Ralph Solecki to claim that the man buried was some leader, healer, or shaman, and that "The association of flowers with Neanderthals adds a whole new dimension to our knowledge of his humanness, indicating that he had 'soul' ".[347] However, it is also possible the pollen was deposited by a small rodent after the man's death.[348]
The graves of children and infants, especially, are associated with grave goods such as artefacts and bones. The grave of a newborn from La Ferrassie, France, was found with three flint scrapers, and an infant from Cave, Syria, was found with a triangular flint placed on its chest. A 10-month-old from Amud Cave, Israel, was associated with a red deer mandible, likely purposefully placed there given other animal remains are now reduced to fragments. Teshik-Tash 1 from Uzbekistan was associated with a circle of ibex horns, and a limestone slab argued to have supported the head.[227] A child from Kiik-Koba, Crimea, Ukraine, had a flint flake with some purposeful engraving on it, likely requiring a great deal of skill.[55] Nonetheless, these contentiously constitute evidence of symbolic meaning as the grave goods' significance and worth are unclear.[227]
Cults[]
It was once argued that the bones of the cave bear, particularly the skull, in some European caves were arranged in a specific order, indicating an ancient bear cult that killed bears and then ceremoniously arranged the bones. This would be consistent with bear-related rituals of modern human Arctic hunter-gatherers, but the alleged peculiarity of the arrangement could also be well-explained by natural causes,[60][342] and bias could be introduced as the existence of a bear cult would conform with the idea that totemism was the earliest religion, leading to undue extrapolation of evidence.[349]
It was also once thought that Neanderthals ritually hunted, killed, and cannibalised other Neanderthals and used the skull as the focus of some ceremony.[267] In 1962, Italian palaeontologist Alberto Blanc believed a skull from Grotta Guattari, Italy, had evidence of a swift blow to the head—indicative of ritual murder—and a precise and deliberate incising at the base to access the brain. He compared it to the victims of headhunters in Malaysia and Borneo,[350] putting it forward as evidence of a skull cult.[342] However, it is now thought to have been a result of cave hyaena predation.[351] Although Neanderthals are known to have practiced cannibalism, there is unsubstantial evidence to suggest ritual defleshing.[266]
In 2019, Gibraltarian palaeoanthropologists Stewart, Geraldine, and Clive Finlayson and Spanish archaeologist Francisco Guzmán speculated that the golden eagle had iconic value to Neanderthals, as exemplified in some modern human societies because they reported that golden eagle bones had a conspicuously high rate of evidence of modification compared to the bones of other birds. They then proposed some "Cult of the Sun Bird" where the golden eagle was a symbol of power.[280][52]
Interbreeding[]
Interbreeding with modern humans[]

The first Neanderthal genome sequence was published in 2010, and strongly indicated interbreeding between Neanderthals and early modern humans.[76][353][354][355] The genomes of all non-sub-Saharan populations contain Neanderthal DNA.[76][78][356][357] Various estimates exist for the proportion, such as 1–4%[76] or 3.4–7.9% in modern Eurasians,[358] or 1.8–2.4% in modern Europeans and 2.3–2.6% in modern East Asians.[359] Pre-agricultural Europeans appear to have had similar percentages to modern East Asians, and the numbers may have decreased in the former due to dilution with a group of people which had split off before Neanderthal introgression.[89] Typically, studies have reported finding no significant levels of Neanderthal DNA in Subsaharan Africans, but a 2020 study detected 0.3-0.5% in the genomes of five African sample populations, likely the result of Eurasians back-migrating and interbreeding with Africans, as well as human-to-neanderthal gene flow from dispersals of Homo sapiens preceding the larger Out-of-Africa migration.[360] Such low percentages of Neanderthal DNA in all present day populations indicate infrequent past interbreeding,[361] unless interbreeding was more common with a different population of modern humans which did not contribute to the present day gene pool.[89] Of the inherited Neanderthal genome, 25% in modern Europeans and 32% in modern East Asians may be related to viral immunity.[362] In all, approximately 20% of the Neanderthal genome appears to have survived in the modern human gene pool.[83]
However, due to their small population and resulting reduced effectivity of natural selection, Neanderthals accumulated several weakly harmful mutations, which were introduced to and slowly selected out of the much larger modern human population; the initial hybridised population may have experienced up to a 94% reduction in fitness compared to contemporary humans. By this measure, Neanderthals may have substantially increased in fitness.[71] A 2017 study focusing on archaic genes in Turkey found associations with coeliac disease, malaria severity, and Costello syndrome.[363] Nonetheless, some genes may have helped modern human Europeans adapt to the environment; the Val92Met variant of the MC1R gene, which may be weakly associated with red hair,[364] may descend from Neanderthals[365] although this is contested as the variant was rare in Neanderthals,[198] and light skin in modern humans did not become prevalent until the Holocene.[199] Some genes related to the immune system appear to have been affected by introgression, which may have aided migration,[85] such as OAS1,[366] STAT2,[367] TLR6, TLR1, TLR10,[368] and several related to immune response.[84][f] In addition, Neanderthal genes have also been implicated in the structure and function of the brain,[g] keratin filaments, sugar metabolism, muscle contraction, body fat distribution, enamel thickness, and oocyte meiosis.[88] Nonetheless, a large portion of surviving introgression appears to be non-coding ("junk") DNA with few biological functions.[89]
Due to the absence of Neanderthal-derived mtDNA (which is passed on from mother to child) in modern populations,[124][141][370] it has been suggested that the progeny of Neanderthal females who mated with modern human males were either rare, absent, or sterile—that is to say, admixture stems from the progeny of Neanderthal males with modern human females.[77][122][370][371][89] Due to the lack of Neanderthal-derived Y-chromosomes in modern humans (which is passed on from father to son), it has also been suggested that the hybrids that contributed ancestry to modern populations were predominantly females, or the Neanderthal Y-chromosome was not compatible with H. sapiens and became extinct.[372][89]
According to linkage disequilibrium mapping, the last Neanderthal gene flow into the modern human genome occurred 86–37 thousand years ago, but most likely 65–47 thousand years ago.[373] It is thought that Neanderthal genes which contributed to the present day human genome stemmed from interbreeding in the Near East rather than the entirety of Europe. However, interbreeding still occurred without contributing to the modern genome[89] The approximately 40,000 year old modern human Oase 2 was found, in 2015, to have had 6–9% (point estimate 7.3%) Neanderthal DNA, indicating a Neanderthal ancestor up to four to six generations earlier, but this hybrid Romanian population does not appear to have made a substantial contribution to the genomes of later Europeans.[352] In 2016, the DNA of Neanderthals from Denisova Cave revealed evidence of interbreeding 100,000 years ago, and interbreeding with an earlier dispersal of H. sapiens may have occurred as early as 120,000 years ago in places such as the Levant.[80] The earliest H. sapiens remains outside of Africa occur at Misliya Cave 194–177 thousand years ago, and Skhul and Qafzeh 120–90 thousand years ago.[374] The Qafzeh humans lived at approximately the same time as the Neanderthals from the nearby Tabun Cave.[375] The Neanderthals of the German Hohlenstein-Stadel have deeply divergent mtDNA compared to more recent Neanderthals, possibly due to introgression of human mtDNA between 316 and 219 thousand years ago, or simply because they were genetically isolated.[79] Whatever the case, these first interbreeding events have not left any trace in modern human genomes.[376]
Detractors of the interbreeding model argue that the genetic similarity is only a remnant of a common ancestor instead of interbreeding,[377] although this is unlikely as it fails to explain why sub-Saharan Africans do not have Neanderthal DNA.[354]
Interbreeding with Denisovans[]

Although nDNA confirms that Neanderthals and Denisovans are more closely related to each other than they are to modern humans, Neanderthals and modern humans share a more recent maternally-transmitted mtDNA common ancestor, possibly due to interbreeding between Denisovans and some unknown human species. The 400,000-year-old Neanderthal-like humans from Sima de los Huesos in northern Spain, looking at mtDNA, are more closely related to Denisovans than Neanderthals. Several Neanderthal-like fossils in Eurasia from a similar time period are often grouped into H. heidelbergensis, of which some may be relict populations of earlier humans, which could have interbred with Denisovans.[379] This is also used to explain an approximately 124,000 year old German Neanderthal specimen with mtDNA that diverged from other Neanderthals (except for Sima de los Huesos) about 270,000 years ago, while its genomic DNA indicated divergence less than 150,000 years ago.[79]
Sequencing of the genome of a Denisovan from Denisova Cave has shown that 17% of its genome derives from Neanderthals.[82] This Neanderthal DNA more closely resembled that of a 120,000-year-old Neanderthal bone from the same cave than that of Neanderthals from Vindija Cave, Croatia, or Mezmaiskaya Cave in the Caucasus, suggesting that interbreeding was local.[81]
For the 90,000-year-old Denisova 11, it was found that her father was a Denisovan related to more recent inhabitants of the region, and her mother a Neanderthal related to more recent European Neanderthals at Vindija Cave, Croatia. Given how few Denisovan bones are known, the discovery of a first-generation hybrid indicates interbreeding was very common between these species, and Neanderthal migration across Eurasia likely occurred sometime after 120,000 years ago.[380]
Extinction[]
Transition[]
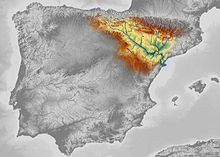
Whatever the cause of their extinction, Neanderthals were replaced by modern humans, indicated by near full replacement of Middle Palaeolithic Mousterian stone technology with modern human Upper Palaeolithic Aurignacian stone technology across Europe (the Middle-to-Upper Palaeolithic Transition) from 41 to 39 thousand years ago.[9][10][12][381] However, it is postulated that Iberian Neanderthals persisted until about 35,000 years ago indicated by the date range of transitional lithic assemblages—Châtelperronian, Uluzzian, Protoaurignacian, and Early Aurignacian. The latter two are attributed to modern humans, but the former two have unconfirmed authorship, potentially products of Neanderthal/modern human cohabitation and cultural transmission. Further, the appearance of the Aurignacian south of the Ebro River has been dated to roughly 37,500 years ago, which has prompted the "Ebro Frontier" hypothesis which states that the river presented a geographic barrier preventing modern human immigration, and thus prolonging Neanderthal persistence.[382][383] However, the dating of the Iberian Transition is debated, with a contested timing of 43–40.8 thousand years ago at Cueva Bajondillo, Spain.[384][385][386][387] The Châtelperronian appears in northeastern Iberia about 42.5–41.6 thousand years ago.[382]
Some Neanderthals in Gibraltar were dated to much later than this—such as Zafarraya (30,000 years ago)[388] and Gorham's Cave (28,000 years ago)[389]—which may be inaccurate as they were based on ambiguous artefacts instead of direct dating.[12] A claim of Neanderthals surviving in a polar refuge in the Ural Mountains[154] is loosely supported by Mousterian stone tools dating to 34–31 thousand years ago from the northern Siberian site at a time when modern humans may not yet have colonised the northern reaches of Europe;[156] however, modern human remains are known from the nearby Mamontovaya Kurya site dating to 40,000 years ago.[390] Indirect dating of Neanderthals remains from Mezmaiskaya Cave reported a date of about 30,000 years ago, but direct dating instead yielded 39.7±1.1 thousand years ago, more in line with trends exhibited in the rest of Europe.[11]

The earliest indication of Upper Palaeolithic modern human immigration into Europe is the Balkan Bohunician industry beginning 48,000 years ago, likely deriving from the Levantine Emiran industry,[313] and the earliest bones in Europe date to roughly 45–43 thousand years ago in Bulgaria,[391] Italy,[392] and Britain.[393] This wave of modern humans replaced Neanderthals.[9] However, Neanderthals and H. sapiens have a much longer contact history. DNA evidence indicates H. sapiens contact with Neanderthals and admixture as early as 120–100 thousand years ago. A 2019 reanalysis of 210,000 year old skull fragments from the Greek Apidima Cave assumed to have belonged to a Neanderthal concluded that they belonged to a modern human, and a Neanderthal skull dating to 170,000 years ago from the cave indicates H. sapiens were replaced by Neanderthals until returning about 40,000 years ago.[394] This identification was refuted by a 2020 study.[395] Archaeological evidence suggests that Neanderthals displaced modern humans in the Near East around 100,000 years ago until about 60–50 thousand years ago.[89]
Cause[]
Modern humans[]
Historically, modern human technology was viewed as vastly superior to that of Neanderthals, with more efficient weaponry and subsistence strategies, and Neanderthals simply went extinct because they could not compete.[13]
The discovery of Neanderthal/modern human introgression has caused the resurgence of the multiregional hypothesis, wherein the present day genetic makeup of all humans is the end result of complex genetic contact among several different populations of humans dispersed across the world. By this model, Neanderthals and other recent archaic humans were simply assimilated into the modern human genome – that is, they were effectively bred out into extinction.[13]
Climate change[]
Their ultimate extinction coincides with Heinrich event 4, a period of intense seasonality; later Heinrich events are also associated with massive cultural turnovers when European human populations collapsed.[14][15] This climate change may have depopulated several regions of Neanderthals, like previous cold spikes, but these areas were instead repopulated by immigrating humans, leading to Neanderthal extinction.[396]

It has also been proposed that climate change was the primary driver, as their low population left them vulnerable to any environmental change, with even a small drop in survival or fertility rates possibly quickly leading to their extinction.[397] However, Neanderthals and their ancestors had survived through several glacial periods over their hundreds of thousands of years of European habitation.[247] It is also proposed that around 40,000 years ago, when Neanderthal populations may have already been dwindling from other factors, the Campanian Ignimbrite Eruption in Italy could have led to their final demise, as it produced 2–4 °C cooling for a year and acid rain for several more years.[16][398]
Disease[]
Modern humans may have introduced African diseases to Neanderthals, contributing to their extinction. A lack of immunity, compounded by an already low population, was potentially devastating to the Neanderthal population, and low genetic diversity could have also rendered fewer Neanderthals naturally immune to these new diseases ("differential pathogen resistance" hypothesis). However, compared to modern humans, Neanderthals had a similar or higher genetic diversity for 12 major histocompatibility complex (MHC) genes associated with the adaptive immune system, casting doubt on this model.[18]
Low population and inbreeding depression may have caused maladaptive birth defects, which could have contributed to their decline (mutational meltdown).[217]
In late-20th-century New Guinea, due to cannibalistic funerary practices, the Fore people were decimated by transmissible spongiform encephalopathies, specifically kuru, a highly virulent disease spread by ingestion of prions found in brain tissue. However, individuals with the 129 variant of the PRNP gene were naturally immune to the prions. Studying this gene led to the discovery that the 129 variant was widespread among all modern humans, which could indicate widespread cannibalism at some point in human prehistory. Because Neanderthals are known to have practised cannibalism to an extent and to have co-existed with modern humans, British palaeoanthropologist Simon Underdown speculated that modern humans transmitted a kuru-like spongiform disease to Neanderthals, and, because the 129 variant appears to have been absent in Neanderthals, it quickly killed them off.[17][399]
In popular culture[]

Neanderthals have been portrayed in popular culture including appearances in literature, visual media, and comedy. The "caveman" archetype often mocks Neanderthals and depicts them as primitive, hunchbacked, knuckle-dragging, club-wielding, grunting, nonsocial characters driven solely by animal instinct. "Neanderthal" can also be used as an insult.[23]
In literature, they are sometimes depicted as brutish or monstrous, such as in H. G. Wells' The Grisly Folk and Elizabeth Marshall Thomas' The Animal Wife, but sometimes with a civilised but unfamiliar culture, as in William Golding's The Inheritors, Björn Kurtén's Dance of the Tiger, and Jean M. Auel's Clan of the Cave Bear and her Earth's Children series.[24]
See also[]
- Denisovan – Asian archaic human
- Early human migrations
- European early modern humans
- Homo floresiensis – Archaic human from Flores, Indonesia
- Homo luzonensis – Archaic human from Luzon, Philippines
- Homo naledi – Small-brained South African archaic human
- Human timeline – Hominin events for the last 10 million years
Explanatory notes[]
- ^ After being mined for limestone, the cave caved in and was lost by 1900. It was rediscovered in 1997 by archaeologists and .[90]
- ^ The German spelling Thal ("valley") was current until 1901 but has been Tal since then. (The German noun is cognate with English dale.) The German /t/ phoneme was frequently spelled th from the 15th to 19th centuries, but the spelling Tal became standardized with the German spelling reform of 1901 and the old spellings of the German names Neanderthal for the valley and Neanderthaler for the species were both changed to the spellings without h.[91][92]
- ^ In Mettmann, "Neander Valley", there is a local idiosyncrasy in use of the outdated spellings with th, such as with the Neanderthal Museum (but the name is in English [German would require Neandertalermuseum]), the Neanderthal station (Bahnhof Neanderthal), and some other rare occasions meant for tourists. Beyond these, city convention is to use th when referring to the species.[92]
- ^ King made a typo and said "theositic".
- ^ The bones were discovered by workers of and . Initially, the workers threw the bones out as debris, but Beckershoff then told them to store the bones. Pieper asked Fuhlrott to come up to the cave and investigate the bones, which Beckershoff and Pieper believed belonged to a cave bear.[90]
- ^ OAS1[366] and STAT2[367] both are associated with fighting viral inflections (interferons), and the listed toll-like receptors (TLRs)[368] allow cells to identify bacterial, fungal, or parasitic pathogens. African origin is also correlated with a stronger inflammatory response.[84]
- ^ Higher levels of Neanderthal-derived genes are associated with an occipital and parietal bone shape reminiscent to that of Neanderthals, as well as modifications to the visual cortex and the intraparietal sulcus (associated with visual processing).[369]
- ^ Homo floresiensis originated in an unknown location from unknown ancestors and reached remote parts of Indonesia. Homo erectus spread from Africa to western Asia, then east Asia and Indonesia; its presence in Europe is uncertain, but it gave rise to Homo antecessor, found in Spain. Homo heidelbergensis originated from Homo erectus in an unknown location and dispersed across Africa, southern Asia and southern Europe (other scientists interpret fossils, here named heidelbergensis, as late erectus). Modern humans spread from Africa to western Asia and then to Europe and southern Asia, eventually reaching Australia and the Americas. In addition to Neanderthals and Denisovans, a third gene flow of archaic Africa origin is indicated at the right.[378] The chart is missing superarchaic (which diverged from erectus 1.9 mya) introgression into Neanderthal/Denisovan common ancestor.[145]
Citations[]
- ^ a b Haeckel, E. (1895). Systematische Phylogenie: Wirbelthiere (in German). G. Reimer. p. 601.
- ^ Schwalbe, G. (1906). Studien zur Vorgeschichte des Menschen [Studies on the pre-history of man] (in German). Stuttgart, E. Nägele. doi:10.5962/bhl.title.61918. hdl:2027/uc1.b4298459.
- ^ Klaatsch, H. (1909). "Preuves que l'Homo Mousteriensis Hauseri appartient au type de Neandertal" [Evidence that Homo Mousteriensis Hauseri belongs to the Neanderthal type]. L'Homme Préhistorique (in French). 7: 10–16.
- ^ Romeo, L. (1979). Ecce Homo!: a lexicon of man. John Benjamins Publishing Company. p. 92. ISBN 978-90-272-2006-6.
- ^ a b c d e McCown, T.; Keith, A. (1939). The stone age of Mount Carmel. The fossil human remains from the Levalloisso-Mousterian. 2. Clarenden Press.
- ^ Szalay, F. S.; Delson, E. (2013). Evolutionary history of the Primates. Academic Press. p. 508. ISBN 978-1-4832-8925-0.
- ^ Wells, J. (2008). Longman pronunciation dictionary (3rd ed.). Harlow, England: Pearson Longman. ISBN 978-1-4058-8118-0.
- ^ a b c d Pääbo, S. (2014). Neanderthal man: in search of lost genomes. New York: Basic Books. p. 237.
- ^ a b c Higham, T.; Douka, K.; Wood, R.; Ramsey, C. B.; Brock, F.; Basell, L.; Camps, M.; Arrizabalaga, A.; Baena, J.; Barroso-Ruíz, C.; C. Bergman; C. Boitard; P. Boscato; M. Caparrós; N.J. Conard; C. Draily; A. Froment; B. Galván; P. Gambassini; A. Garcia-Moreno; S. Grimaldi; P. Haesaerts; B. Holt; M.-J. Iriarte-Chiapusso; A. Jelinek; J.F. Jordá Pardo; J.-M. Maíllo-Fernández; A. Marom; J. Maroto; M. Menéndez; L. Metz; E. Morin; A. Moroni; F. Negrino; E. Panagopoulou; M. Peresani; S. Pirson; M. de la Rasilla; J. Riel-Salvatore; A. Ronchitelli; D. Santamaria; P. Semal; L. Slimak; J. Soler; N. Soler; A. Villaluenga; R. Pinhasi; R. Jacobi; et al. (2014). "The timing and spatiotemporal patterning of Neanderthal disappearance". Nature. 512 (7514): 306–309. Bibcode:2014Natur.512..306H. doi:10.1038/nature13621. hdl:1885/75138. PMID 25143113. S2CID 205239973.
We show that the Mousterian [the Neanderthal tool-making tradition] ended by 41,030–39,260 calibrated years BP (at 95.4% probability) across Europe. We also demonstrate that succeeding 'transitional' archaeological industries, one of which has been linked with Neanderthals (Châtelperronian), end at a similar time.
- ^ a b Higham, T. (2011). "European Middle and Upper Palaeolithic radiocarbon dates are often older than they look: problems with previous dates and some remedies". Antiquity. 85 (327): 235–249. doi:10.1017/s0003598x00067570. S2CID 163207571.
Few events of European prehistory are more important than the transition from ancient to modern humans about 40,000 years ago, a period that unfortunately lies near the limit of radiocarbon dating. This paper shows that as many as 70 per cent of the oldest radiocarbon dates in the literature may be too young, due to contamination by modern carbon.
- ^ a b Pinhasi, R.; Higham, T. F. G.; Golovanova, L. V.; Doronichev, V. B. (2011). "Revised age of late Neanderthal occupation and the end of the Middle Palaeolithic in the northern Caucasus". Proceedings of the National Academy of Sciences. 108 (21): 8611–8616. Bibcode:2011PNAS..108.8611P. doi:10.1073/pnas.1018938108. PMC 3102382. PMID 21555570.
The direct date of the fossil (39,700 ± 1,100 14C BP) is in good agreement with the probability distribution function, indicating at a high level of probability that Neanderthals did not survive at Mezmaiskaya Cave after 39 kya cal BP. [...] This challenges previous claims for late Neanderthal survival in the northern Caucasus. [...] Our results confirm the lack of reliably dated Neanderthal fossils younger than ≈40 kya cal BP in any other region of Western Eurasia, including the Caucasus.
- ^ a b c Galván, B.; Hernández, C. M.; Mallol, C.; Mercier, N.; Sistiaga, A.; Soler, V. (2014). "New evidence of early Neanderthal disappearance in the Iberian Peninsula". Journal of Human Evolution. 75: 16–27. doi:10.1016/j.jhevol.2014.06.002. PMID 25016565.
- ^ a b c Villa, P.; Roebroeks, W. (2014). "Neandertal demise: an archaeological analysis of the modern human superiority complex". PLOS ONE. 9 (4): e96424. Bibcode:2014PLoSO...996424V. doi:10.1371/journal.pone.0096424. PMC 4005592. PMID 24789039.
- ^ a b Bradtmöller, M.; Pastoors, A.; Weninger, B.; Weninger, G. (2012). "The repeated replacement model – Rapid climate change and population dynamics in Late Pleistocene Europe". Quaternary International. 247: 38–49. Bibcode:2012QuInt.247...38B. doi:10.1016/j.quaint.2010.10.015.
- ^ a b Wolf, D.; Kolb, T.; Alcaraz-Castaño, M.; Heinrich, S. (2018). "Climate deteriorations and Neanderthal demise in interior Iberia". Scientific Reports. 8 (1): 7048. Bibcode:2018NatSR...8.7048W. doi:10.1038/s41598-018-25343-6. PMC 5935692. PMID 29728579.
- ^ a b c Black, B. A.; Neely, R. R.; Manga, M. (2015). "Campanian Ignimbrite volcanism, climate, and the final decline of the Neanderthals" (PDF). Geology. 43 (5): 411–414. Bibcode:2015Geo....43..411B. doi:10.1130/G36514.1. OSTI 1512181.
- ^ a b Underdown, S. (2008). "A potential role for transmissible spongiform encephalopathies in Neanderthal extinction". Medical Hypotheses. 71 (1): 4–7. doi:10.1016/j.mehy.2007.12.014. PMID 18280671.
- ^ a b Sullivan, A. P.; de Manuel, M.; Marques-Bonet, T.; Perry, G. H. (2017). "An evolutionary medicine perspective on Neandertal extinction" (PDF). Journal of Human Evolution. 108: 62–71. doi:10.1016/j.jhevol.2017.03.004. PMID 28622932.
- ^ a b c Stringer, C. (2012). "The status of Homo heidelbergensis (Schoetensack 1908)". Evolutionary Anthropology. 21 (3): 101–107. doi:10.1002/evan.21311. PMID 22718477. S2CID 205826399.
- ^ a b Gómez-Robles, A. (2019). "Dental evolutionary rates and its implications for the Neanderthal–modern human divergence". Science Advances. 5 (5): eaaw1268. Bibcode:2019SciA....5.1268G. doi:10.1126/sciadv.aaw1268. PMC 6520022. PMID 31106274.
- ^ a b c d e f Meyer, M.; Arsuaga, J.; de Filippo, C.; Nagel, S. (2016). "Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins". Nature. 531 (7595): 504–507. Bibcode:2016Natur.531..504M. doi:10.1038/nature17405. PMID 26976447. S2CID 4467094.
- ^ a b Klein, R. G. (1983). "What Do We Know About Neanderthals and Cro-Magnon Man?". Anthropology. 52 (3): 386–392. JSTOR 41210959.
- ^ a b c d e f g Papagianni & Morse 2013.
- ^ a b c d e f g h i Drell, J. R. R. (2000). "Neanderthals: a history of interpretation". Oxford Journal of Archaeology. 19 (1): 1–24. doi:10.1111/1468-0092.00096. S2CID 54616107.
- ^ a b Shaw, I.; Jameson, R., eds. (1999). A dictionary of archaeology. Blackwell. p. 408. ISBN 978-0-631-17423-3.
- ^ a b Lycett, S. J.; von Cramon-Taubadel, N. (2013). "A 3D morphometric analysis of surface geometry in Levallois cores: patterns of stability and variability across regions and their implications". Journal of Archaeological Science. 40 (3): 1508–1517. doi:10.1016/j.jas.2012.11.005.
- ^ a b Sorensen, A. C.; Claud, E.; Soressi, M. (2018). "Neandertal fire-making technology inferred from microwear analysis". Scientific Reports. 8 (1): 10065. Bibcode:2018NatSR...810065S. doi:10.1038/s41598-018-28342-9. ISSN 2045-2322. PMC 6053370. PMID 30026576.
- ^ a b Brittingham, A.; Hren, M. T.; Hartman, G.; Wilkinson, K. N.; Mallol, C.; Gasparyan, B.; Adler, D. S. (2019). "Geochemical evidence for the control of fire by Middle Palaeolithic hominins". Scientific Reports. 9 (1): 15368. Bibcode:2019NatSR...915368B. doi:10.1038/s41598-019-51433-0. PMC 6814844. PMID 31653870.
- ^ a b c d e f g h i j k Hayden, B. (2012). "Neandertal social structure?". Oxford Journal of Archaeology. 31 (1): 1–26. doi:10.1111/j.1468-0092.2011.00376.x.
- ^ a b c d Kedar, Y.; Barkai, R. (2019). "The significance of air circulation and hearth location at Paleolithic cave sites". Open Quaternary. 5 (1): 4. doi:10.5334/oq.52.
- ^ a b Schmidt, P.; Blessing, M.; Rageot, M.; Iovita, R.; Pfleging, J.; Nickel, K. G.; Righetti, L.; Tennie, C. (2019). "Birch tar production does not prove Neanderthal behavioral complexity". Proceedings of the National Academy of Sciences. 116 (36): 17707–17711. doi:10.1073/pnas.1911137116. PMC 6731756. PMID 31427508.
- ^ a b Hoffecker, J. F. (2009). "The spread of modern humans in Europe". Proceedings of the National Academy of Sciences. 106 (38): 16040–16045. Bibcode:2009PNAS..10616040H. doi:10.1073/pnas.0903446106. PMC 2752585. PMID 19571003.
- ^ a b Hardy, B. L.; Moncel, M.-H.; Kerfant, C.; et al. (2020). "Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications". Scientific Reports. 10 (4889): 4889. Bibcode:2020NatSR..10.4889H. doi:10.1038/s41598-020-61839-w. PMC 7145842. PMID 32273518.
- ^ a b Ferentinos, G.; Gkioni, M.; Geraga, M.; Papatheodorou, G. (2012). "Early seafaring activity in the southern Ionian Islands, Mediterranean Sea". Journal of Archaeological Science. 39 (7): 2167–2176. Bibcode:2011JQS....26..553S. doi:10.1016/j.jas.2012.01.032.
- ^ a b c d Strasser, T. F.; Runnels, C.; Wegmann, K. W.; Panagopoulou, E. (2011). "Dating Palaeolithic sites in southwestern Crete, Greece". Journal of Quaternary Science. 26 (5): 553–560. Bibcode:2011JQS....26..553S. doi:10.1016/j.jas.2012.01.032.
- ^ a b c Buckley, S.; Hardy, K.; Huffman, M. (2013). "Neanderthal self-medication in context". Antiquity. 87 (337): 873–878. doi:10.1017/S0003598X00049528. S2CID 160563162.
- ^ a b c Lev, E.; Kislev, M. E.; Bar-Yosef, O. (2005). "Mousterian vegetal food in Kebara Cave, Mt. Carmel". Journal of Archaeological Science. 32 (3): 475–484. doi:10.1016/j.jas.2004.11.006.
- ^ Power, R. C.; Salazar-García, D. C.; Rubini, M.; Darlas, A.; Harvati, K.; Walker, M.; Hublin, J.; Henry, A. G. (2018). "Dental calculus indicates widespread plant use within the stable Neanderthal dietary niche". Journal of Human Evolution. 119: 27–41. doi:10.1016/j.jhevol.2018.02.009. hdl:10550/65536. ISSN 0047-2484. PMID 29685752.
- ^ a b c d e f Spikins, P.; Needham, A.; Wright, B. (2019). "Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare". Quaternary Science Reviews. 217: 98–118. Bibcode:2019QSRv..217...98S. doi:10.1016/j.quascirev.2018.08.011.
- ^ a b c Valensi, P.; Michel, V.; et al. (2013). "New data on human behavior from a 160,000 year old Acheulean occupation level at Lazaret cave, south-east France: An archaeozoological approach". Quaternary International. 316: 123–139. Bibcode:2013QuInt.316..123V. doi:10.1016/j.quaint.2013.10.034.
- ^ a b c d Krief, S.; Daujeard, C.; Moncel, M.; Lamon, N.; Reynolds, V. (2015). "Flavouring food: the contribution of chimpanzee behaviour to the understanding of Neanderthal calculus composition and plant use in Neanderthal diets". Antiquity. 89 (344): 464–471. doi:10.15184/aqy.2014.7. S2CID 86646905.
- ^ a b Hardy, K.; Buckley, S.; Collins, M. J.; Estalrrich, A. (2012). "Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus". The Science of Nature. 99 (8): 617–626. Bibcode:2012NW.....99..617H. doi:10.1007/s00114-012-0942-0. PMID 22806252. S2CID 10925552.
- ^ a b c d Dusseldorp, G. L. (2013). "Neanderthals and cave hyenas: co-existence, competition or conflict?" (PDF). In Clark, J. L.; Speth, J. D. (eds.). Zooarchaeology and modern human origins. Vertebrate paleobiology and paleoanthropology. Springer Science+Business Media Dordrecht. pp. 191–208. doi:10.1007/978-94-007-6766-9_12. ISBN 978-94-007-6765-2.
- ^ a b c Richards, M. P.; Pettitt, P. B.; Trinkaus, E.; Smith, F. H.; Paunović, M.; Karavanić, I. (2000). "Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes". Proceedings of the National Academy of Sciences. 97 (13): 7663–7666. Bibcode:2000pnas...97.7663r. doi:10.1073/pnas.120178997. JSTOR 122870. PMC 16602. PMID 10852955.
- ^ a b Henry, A. G.; Brooks, A. S.; Piperno, D. R. (2011). "Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium)". Proceedings of the National Academy of Sciences. 108 (2): 486–491. Bibcode:2011PNAS..108..486H. doi:10.1073/pnas.1016868108. PMC 3021051. PMID 21187393.
- ^ a b Shipley, G. P.; Kindscher, K. (2016). "Evidence for the paleoethnobotany of the Neanderthal: a review of the literature". Scientifica. 2016: 1–12. doi:10.1155/2016/8927654. PMC 5098096. PMID 27843675.
- ^ a b Madella, M.; Jones, M. K.; Goldberg, P.; Goren, Y.; Hovers, E. (2002). "The Exploitation of plant resources by Neanderthals in Amud Cave (Israel): the evidence from phytolith studies". Journal of Archaeological Science. 29 (7): 703–719. doi:10.1006/jasc.2001.0743. S2CID 43217031.
- ^ a b c d e f Brown 2011.
- ^ a b Shipman 2015, pp. 120–143.
- ^ a b c d Tattersall 2015.
- ^ a b d’Errico, F.; Tsvelykh, A. (2017). "A decorated raven bone from the Zaskalnaya VI (Kolosovskaya) Neanderthal site, Crimea". PLOS ONE. 12 (3): e0173435. Bibcode:2017PLoSO..1273435M. doi:10.1371/journal.pone.0173435. PMC 5371307. PMID 28355292.
- ^ a b c Finlayson 2019.
- ^ a b Hoffman, D. L.; Angelucci, D. E.; Villaverde, V.; Zapata, Z.; Zilhão, J. (2018). "Symbolic use of marine shells and mineral pigments by Iberian Neandertals 115,000 years ago". Science Advances. 4 (2): eaar5255. Bibcode:2018SciA....4.5255H. doi:10.1126/sciadv.aar5255. PMC 5833998. PMID 29507889.
- ^ a b c Moncel, M.-H.; Chiotti, L.; Gaillard, C.; Onoratini, G.; Pleurdeau, D. (2012). "Non utilitarian objects in the Palaeolithic: emergence of the sense of precious?". Archaeology, Ethnology & Anthropology of Eurasia. 401: 25–27. doi:10.1016/j.aeae.2012.05.004.
- ^ a b c Majkić, A.; d’Errico, F.; Stepanchuk, V. (2018). "Assessing the significance of Palaeolithic engraved cortexes. A case study from the Mousterian site of Kiik-Koba, Crimea". PLOS ONE. 13 (5): e0195049. Bibcode:2018PLoSO..1395049M. doi:10.1371/journal.pone.0195049. PMC 5931501. PMID 29718916.
- ^ a b Turk, M.; Turk, I.; Dimkaroski, L. (2018). "The Mousterian musical instrument from the Divje Babe I Cave (Slovenia): arguments on the material evidence for Neanderthal musical behaviour". L'Anthropologie. 122 (4): 1–28. doi:10.1016/j.anthro.2018.10.001. S2CID 133682741.
- ^ a b Aubert, M.; Brumm, A.; Huntley, J. (2018). "Early dates for 'Neanderthal cave art' may be wrong". Journal of Human Evolution. 125: 215–217. doi:10.1016/j.jhevol.2018.08.004. PMID 30173883. S2CID 52145541.
- ^ a b Pike, A. W.; Hoffmann, D. L.; Pettitt, P. B.; García-Diez, M.; Zilhão, J. (2017). "Dating Palaeolithic cave art: Why U–Th is the way to go" (PDF). Quaternary International. 432: 41–49. Bibcode:2017QuInt.432...41P. doi:10.1016/j.quaint.2015.12.013.
- ^ a b Hoffmann, D. L.; Standish, C. D.; García-Diez, M.; Pettitt, P. B.; Milton, J. A.; Zilhão, J.; Alcolea-González, J. J.; Cantalejo-Duarte, P.; Collado, H.; de Balbín, R.; Lorblanchet, M.; Ramos-Muñoz, J.; Weniger, G.-C.; Pike, A. W. G. (2018). "U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art". Science. 359 (6378): 912–915. Bibcode:2018Sci...359..912H. doi:10.1126/science.aap7778. PMID 29472483.
- ^ a b Wunn, I. (2000). "Beginning of religion". Numen. 47 (4): 417–452. doi:10.1163/156852700511612.
- ^ a b c Dediu, D.; Levinson, S. C. (2018). "Neanderthal language revisited: not only us". Current Opinion in Behavioral Sciences. 21: 49–55. doi:10.1016/j.cobeha.2018.01.001. hdl:21.11116/0000-0000-1667-4. S2CID 54391128.
- ^ a b D’Anastasio, R.; Wroe, S.; Tuniz, C.; Mancini, L.; Cesana, D. T. (2013). "Micro-biomechanics of the Kebara 2 hyoid and its implications for speech in Neanderthals". PLOS ONE. 8 (12): e82261. Bibcode:2013PLoSO...882261D. doi:10.1371/journal.pone.0082261. PMC 3867335. PMID 24367509.
- ^ a b c d e f g Stewart, J.R.; García-Rodríguez, O.; Knul, M.V.; Sewell, L.; Montgomery, H.; Thomas, M.G.; Diekmann, Y. (2019). "Palaeoecological and genetic evidence for Neanderthal power locomotion as an adaptation to a woodland environment". Quaternary Science Reviews. 217: 310–315. Bibcode:2019QSRv..217..310S. doi:10.1016/j.quascirev.2018.12.023. S2CID 133980969.
- ^ a b Kislev, M.; Barkai, R. (2018). "Neanderthal and woolly mammoth molecular resemblance". Human Biology. 90 (2): 115–128. doi:10.13110/humanbiology.90.2.03. PMID 33951886. S2CID 106401104.
- ^ a b de Azevedo, S.; González, M. F.; Cintas, C.; et al. (2017). "Nasal airflow simulations suggest convergent adaptation in Neanderthals and modern humans". Proceedings of the National Academy of Sciences. 114 (47): 12442–12447. doi:10.1073/pnas.1703790114. PMC 5703271. PMID 29087302.
- ^ a b Rae, T. C.; Koppe, T.; Stringer, C. B. (2011). "The Neanderthal face is not cold adapted". Journal of Human Evolution. 60 (2): 234–239. doi:10.1016/j.jhevol.2010.10.003. PMID 21183202.
- ^ a b c d Helmuth, H. (1998). "Body height, body mass and surface area of the Neanderthals". Zeitschrift für Morphologie und Anthropologie. 82 (1): 1–12. doi:10.1127/zma/82/1998/1. JSTOR 25757530. PMID 9850627.
- ^ a b Stringer, C. (1984). "Human evolution and biological adaptation in the Pleistocene". In Foley, R. (ed.). Hominid evolution and community ecology. Academic Press. ISBN 978-0-12-261920-5.
- ^ a b Holloway, R. L. (1985). "The poor brain of Homo sapiens neanderthalensis: see what you please". In Delson, E. (ed.). Ancestors: The hard evidence. Alan R. Liss. ISBN 978-0-471-84376-4.
- ^ a b c Amano, H.; Kikuchi, T.; Morita, Y.; Kondo, O.; Suzuki, H.; et al. (2015). "Virtual reconstruction of the Neanderthal Amud 1 cranium" (PDF). American Journal of Physical Anthropology. 158 (2): 185–197. doi:10.1002/ajpa.22777. hdl:10261/123419. PMID 26249757.
- ^ a b c d Juric, I.; Aeschbacher, S.; Coop, G. (2016). "The strength of selection against Neanderthal introgression". PLOS Genetics. 12 (11): e1006340. doi:10.1371/journal.pgen.1006340. PMC 5100956. PMID 27824859.
- ^ a b c Ruebens, K. (2013). "Regional behaviour among late Neanderthal groups in Western Europe: A comparative assessment of late Middle Palaeolithic bifacial tool variability". Journal of Human Evolution. 65 (4): 341–362. doi:10.1016/j.jhevol.2013.06.009. PMID 23928352.
- ^ a b c d Fabre, V.; Condemi, S.; Degioanni, A. (2009). "Genetic evidence of geographical groups among Neanderthals". PLOS ONE. 4 (4): e5151. Bibcode:2009PLoSO...4.5151F. doi:10.1371/journal.pone.0005151. PMC 2664900. PMID 19367332.
- ^ a b c Demay, L.; Péan, S.; Patou-Mathis, M. (2012). "Mammoths used as food and building resources by Neanderthals: zooarchaeological study applied to layer 4, Molodova I (Ukraine)" (PDF). Quaternary International. 276–277: 212–226. Bibcode:2012QuInt.276..212D. doi:10.1016/j.quaint.2011.11.019. hdl:2268/190618.
- ^ a b c d e f g Trinkaus, E. (1995). "Neanderthal mortality patterns". Journal of Archaeological Science. 22 (1): 121–142. doi:10.1016/S0305-4403(95)80170-7.
- ^ a b c d e Green, R. E.; Krause, J.; Briggs, A. W.; et al. (2010). "A draft sequence of the Neandertal genome". Science. 328 (5979): 710–722. Bibcode:2010Sci...328..710G. doi:10.1126/science.1188021. PMC 5100745. PMID 20448178.
- ^ a b Sankararaman, S.; Mallick, S.; Patterson, N.; Reich, D. (2016). "The combined landscape of Denisovan and Neanderthal ancestry in present-day humans". Current Biology. 26 (9): 1241–1247. doi:10.1016/j.cub.2016.03.037. PMC 4864120. PMID 27032491.
- ^ a b c d Sankararaman, S.; Mallick, S.; Dannemann, M.; Prüfer, K.; Kelso, J.; Pääbo, S.; Patterson, N.; Reich, D. (2014). "The genomic landscape of Neanderthal ancestry in present-day humans". Nature. 507 (7492): 354–357. Bibcode:2014Natur.507..354S. doi:10.1038/nature12961. PMC 4072735. PMID 24476815.
- ^ a b c d e Peyrégne, S.; Slon, V.; Mafessoni, F.; et al. (2019). "Nuclear DNA from two early Neandertals reveals 80 ka of genetic continuity in Europe". Science Advances. 5 (6): eaaw5873. Bibcode:2019SciA....5.5873P. doi:10.1126/sciadv.aaw5873. PMC 6594762. PMID 31249872.
- ^ a b Kuhlwilm, M. (2016). "Ancient gene flow from early modern humans into eastern Neanderthals". Nature. 530 (7591): 429–433. Bibcode:2016Natur.530..429K. doi:10.1038/nature16544. PMC 4933530. PMID 26886800.
- ^ a b c d e f g Prüfer, K.; et al. (2014). "The complete genome sequence of a Neanderthal from the Altai Mountains". Nature. 505 (7481): 43–49. Bibcode:2014Natur.505...43P. doi:10.1038/nature12886. PMC 4031459. PMID 24352235.
- ^ a b c Pennisi, E. (2013). "More genomes from Denisova Cave show mixing of early human groups". Science. 340 (6134): 799. Bibcode:2013Sci...340..799P. doi:10.1126/science.340.6134.799. PMID 23687020.
- ^ a b Vernot, B.; Akey, J. M. (2014). "Resurrecting surviving Neandertal lineages from modern human genomes". Science. 343 (6174): 1017–1021. Bibcode:2014Sci...343.1017V. doi:10.1126/science.1245938. PMID 24476670. S2CID 23003860.
- ^ a b c Nédélec, Y.; Sanz, J.; Baharian, G.; et al. (2016). "Genetic ancestry and natural selection drive population differences in immune responses to pathogens". Cell. 167 (3): 657–669. doi:10.1016/j.cell.2016.09.025. PMID 27768889.
- ^ a b Ségurel, L.; Quintana-Murci, L. (2014). "Preserving immune diversity through ancient inheritance and admixture". Current Opinion in Immunology. 30: 79–84. doi:10.1016/j.coi.2014.08.002. PMID 25190608.
- ^ Zeberg, Hugo; Pääbo, Svante (March 2, 2021). "A genomic region associated with protection against severe COVID-19 is inherited from Neandertals". Proceedings of the National Academy of Sciences. 118 (9): e2026309118. doi:10.1073/pnas.2026309118. ISSN 0027-8424. PMC 7936282. PMID 33593941. S2CID 231943450.
- ^ Zeberg, Hugo; Pääbo, Svante (September 30, 2020). "The major genetic risk factor for severe COVID-19 is inherited from Neanderthals". Nature. 587 (7835): 610–612. Bibcode:2020Natur.587..610Z. doi:10.1038/s41586-020-2818-3. ISSN 1476-4687. PMID 32998156. S2CID 222148977.
- ^ a b Dolgova, O.; Lao, O. (2018). "Evolutionary and medical consequences of archaic introgression into modern human genomes". Genes. 9 (7): 358. doi:10.3390/genes9070358. PMC 6070777. PMID 30022013.
- ^ a b c d e f g h i j k l Reich 2018.
- ^ a b c Schmitz, R. W.; Serre, D.; Bonani, G.; et al. (2002). "The Neandertal type site revisited: interdisciplinary investigations of skeletal remains from the Neander Valley, Germany". Proceedings of the National Academy of Sciences. 99 (20): 13342–13347. Bibcode:2002PNAS...9913342S. doi:10.1073/pnas.192464099. PMC 130635. PMID 12232049.
- ^ Howell, F. C. (1957). "The evolutionary significance of variation and varieties of 'Neanderthal' man". The Quarterly Review of Biology. 32 (4): 330–347. doi:10.1086/401978. JSTOR 2816956. PMID 13506025. S2CID 10857962.
- ^ a b "Neandertal oder Neanderthal? Was ist denn nun richtig?" [Neandertal or Neanderthal? So which is actually right?]. Kreisstadt Mettmann. Retrieved February 1, 2017.
Heute sollten Ortsbezeichnungen das 'Neandertal' ohne 'h' bezeichnen. Alle Namen, die sich auf den prähistorischen Menschen beziehen, führen das 'h'. (Nowadays, place names should refer to the Neander Valley ['Neandertal'] without an 'h'. All names referring to the prehistoric humans have the 'h'.)
- ^ "Neanderthal". Collins English Dictionary. Retrieved February 18, 2020.
- ^ "Neanderthal". Merriam-Webster Dictionary. Retrieved February 18, 2020.
- ^ "Neanderthal". American Heritage Dictionary. Retrieved February 18, 2020.
- ^ Vogt, K. C. (1864). Lectures on man: his place in creation, and in the history of the earth. London, UK: Longman, Green, Longman and Roberts. pp. 302, 473.
- ^ King, W. (1864). "On the Neanderthal skull, or reasons for believing it to belong to the Clydian Period and to a species different from that represented by man". Report of the British Association for the Advancement of Science, Notices and Abstracts, Newcastle-upon-Tyne, 1863. 33: 81–82 – via Biodiversity Heritage Library.
- ^ Murray, J.; Nasheuer, H. P.; Seoighe, C.; McCormack, G. P.; Williams, D. M.; Harper, D. A. T. (2015). "The contribution of William King to the early development of palaeoanthropology". Irish Journal of Earth Sciences. 33: 1–16. doi:10.3318/ijes.2015.33.1. JSTOR 10.3318/ijes.2015.33.1.
- ^ Winner, A. K. (1964). "Terminology". Current Anthropology. 5 (2): 119–122. doi:10.1086/200469. JSTOR 2739959. S2CID 224796921.
- ^ a b c King, W. (1864). "The reputed fossil man of the Neanderthal" (PDF). Quarterly Journal of Science. 1: 96.
- ^ Schmerling, P. (1834). Recherches sur les ossemens fossiles découverts dans les cavernes de la province de Liége [Research on the fossil specimens discovered in the caves of Liège]. P. J. Collardin. pp. 30–32. hdl:2268/207986.
- ^ Menez, A. (2018). "Custodian of the Gibraltar skull: the history of the Gibraltar Scientific Society". Earth Sciences History. 37 (1): 34–62. doi:10.17704/1944-6178-37.1.34.
- ^ Schaaffhausen, H. (1858). "Zur Kenntnis der ältesten Rassenschädel" [Acknowledging the oldest racial skull]. Archiv für Anatomie, Physiologie und Wissenschaftliche Medicin (in German): 453–478.
- ^ a b c d Schlager, S.; Wittwer-Backofen, U. (2015). "Images in paleoanthropology: facing our ancestors". In Henke, W.; Tattersall, I. (eds.). Handbook of paleoanthropology. Springer-Verlag Berlin Heidelberg. pp. 1019–1027. doi:10.1007/978-3-642-39979-4_70. ISBN 978-3-642-39978-7.
- ^ Fuhlrott, J. C. (1859). "Menschliche Überreste aus einer Felsengrotte des Düsselthales" [Human remains from a rock grotto in Düsseltal] (PDF). Verh Naturhist Ver Preuss Rheinl (in German). 16: 131–153.
- ^ Virchow, R. (1872). "Untersuchung des Neanderthal-Schädels" [Examinations on the Neandertal skull]. Verh Berl Anthrop Ges (in German). 4: 157–165.
- ^ Boule, M. (1911). L'homme fossile de La Chapelle-aux-Saints [Fossil man from La Chapelle-aux-Saints] (in French). Masson. pp. 1–62.
- ^ Van Reybrouck, D. (2002). "Boule's error: on the social context of scientific knowledge". Antiquity. 76 (291): 158–164. doi:10.1017/S0003598X00089936. S2CID 164060946.
- ^ a b c d Langdon, J. H. (2016). "Case study 18. Neanderthals in the mirror: imagining our relatives". The science of human evolution: getting it right. Springer. ISBN 978-3-319-41584-0.
- ^ Sommer, M. (2006). "Mirror, mirror on the wall: Neanderthal as image and 'distortion' in early 20th-century French science and press" (PDF). Social Studies of Science. 36 (2): 207–240. doi:10.1177/0306312706054527. S2CID 145778787.
- ^ Cairney, C. T. (1989). Clans and families of Ireland and Scotland, an ethnography of the Gael. McFarland. p. 14. ISBN 978-0-89950-362-2.
- ^ Huxley, T. (1891). "The Aryan question and pre-historic man". The Popular Science Monthly. 38: 512–516.
- ^ Steensby, H. P. (1907). "Racestudier i Danmark" [Race Studies in Denmark] (PDF). Geographical Journal (in Danish). Geografisk Tidsskrift. 9.
- ^ Coon, C. S. (1962). "The origin of races". Science. Knopf. 140 (3563): 548–549. doi:10.1126/science.140.3563.208. PMID 14022816.
- ^ Tattersall, I.; Schwartz, J. H. (1999). "Hominids and hybrids: The place of Neanderthals in human evolution". Proceedings of the National Academy of Sciences. 96 (13): 7117–19. Bibcode:1999PNAS...96.7117T. doi:10.1073/pnas.96.13.7117. JSTOR 48019. PMC 33580. PMID 10377375.
- ^ Duarte, C.; Maurício, J.; Pettitt, P. B.; Souto, P.; Trinkaus, E.; van der Plicht, H.; Zilhão, J. (1999). "The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia". Proceedings of the National Academy of Sciences of the United States of America. 96 (13): 7604–7609. Bibcode:1999PNAS...96.7604D. doi:10.1073/pnas.96.13.7604. PMC 22133. PMID 10377462.
- ^ a b Hublin, J. J. (2009). "The origin of Neandertals". Proceedings of the National Academy of Sciences. 106 (38): 16022–16027. Bibcode:2009PNAS..10616022H. doi:10.1073/pnas.0904119106. JSTOR 40485013. PMC 2752594. PMID 19805257.
- ^ Harvati, K.; Frost, S. R.; McNulty, K. P. (2004). "Neanderthal taxonomy reconsidered: implications of 3D primate models of intra- and interspecific differences". Proceedings of the National Academy of Sciences. 101 (5): 1147–52. Bibcode:2004PNAS..101.1147H. doi:10.1073/pnas.0308085100. PMC 337021. PMID 14745010.
- ^ Soficaru, A.; Dobos, A.; Trinkaus, E. (2006). "Early modern humans from the Peștera Muierii, Baia de Fier, Romania". Proceedings of the National Academy of Sciences. 103 (46): 17196–17201. Bibcode:2006PNAS..10317196S. doi:10.1073/pnas.0608443103. JSTOR 30052409. PMC 1859909. PMID 17085588.
- ^ a b Chen, F.; Welker, F.; Shen, C. (2019). "A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau" (PDF). Nature. 569 (7, 756): 409–412. Bibcode:2019Natur.569..409C. doi:10.1038/s41586-019-1139-x. PMID 31043746. S2CID 141503768.
- ^ Hofreiter, M. (2011). "Drafting human ancestry: what does the Neanderthal genome tell us about hominid evolution? Commentary on Green et al. (2010)". Human Biology. 83 (1): 1–11. doi:10.3378/027.083.0101. PMID 21453001. S2CID 15005225.
- ^ a b Currat, M.; Excoffier, L. (2004). "Modern humans did not admix with Neanderthals during their range expansion into Europe". PLOS Biology. 2 (12): e421. doi:10.1371/journal.pbio.0020421. PMC 532389. PMID 15562317.
- ^ Mendez, F. L.; Poznik, G. D.; Castellano, S.; Bustamante, C. D. (2016). "The divergence of Neandertal and modern human Y chromosomes". American Journal of Human Genetics. 98 (4): 728–734. doi:10.1016/j.ajhg.2016.02.023. PMC 4833433. PMID 27058445.
- ^ a b Serre, D.; Langaney, A.; Chech, M.; Teschler-Nicola, M.; Paunovic, M.; Mennecier, P.; Hofreiter, M.; Possnert, G.; Pääbo, S. (2004). "No evidence of Neandertal mtDNA contribution to early modern humans". PLOS Biology. 2 (3): e57. doi:10.1371/journal.pbio.0020057. PMC 368159. PMID 15024415.
- ^ a b c d Rogers, A. R.; Bohlender, R. J.; Huff, C. D. (2017). "Early history of Neanderthals and Denisovans". Proceedings of the National Academy of Sciences. 114 (37): 9859–9863. doi:10.1073/pnas.1706426114. PMC 5604018. PMID 28784789.
- ^ a b c Hajdinjak, M.; Fu, Q.; Hübner, A. (2018). "Reconstructing the genetic history of late Neanderthals". Nature. 555 (7698): 652–656. Bibcode:2018Natur.555..652H. doi:10.1038/nature26151. PMC 6485383. PMID 29562232.
- ^ Posth, C.; Wißing, C.; Kitagawa, K.; et al. (2017). "Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals". Nature Communications. 8: 16046. Bibcode:2017NatCo...816046P. doi:10.1038/ncomms16046. PMC 5500885. PMID 28675384.
- ^ a b c Dean, D.; Hublin, J.-J.; Holloway, R.; Ziegler, R. (1998). "On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany". Journal of Human Evolution. 34 (5): 485–508. doi:10.1006/jhev.1998.0214. PMID 9614635. S2CID 6996233.
- ^ Ko, K. W. (2016). "Hominin interbreeding and the evolution of human variation". Journal of Biological Research-Thessaloniki. 23: 17. doi:10.1186/s40709-016-0054-7. PMC 4947341. PMID 27429943.
- ^ Bischoff, J. L.; Shamp, D. D.; Aramburu, A.; et al. (2003). "The Sima de los Huesos hominids date to beyond U/Th equilibrium (>350kyr) and perhaps to 400–500kyr: new radiometric dates". Journal of Archaeological Science. 30 (3): 275–280. doi:10.1006/jasc.2002.0834.
- ^ Arsuaga, J. L.; Martínez, I.; Gracia, A.; Lorenzo, C. (1997). "The Sima de los Huesos crania (Sierra de Atapuerca, Spain). A comparative study". Journal of Human Evolution. 33 (2–3): 219–281. doi:10.1006/jhev.1997.0133. PMID 9300343.
- ^ Daura, J.; Sanz, M.; Arsuaga, J. L.; Hoffman, D. L. (2017). "New Middle Pleistocene hominin cranium from Gruta da Aroeira (Portugal)". Proceedings of the National Academy of Sciences. 114 (13): 3397–3402. doi:10.1073/pnas.1619040114. PMC 5380066. PMID 28289213.
- ^ Stringer 1993, pp. 65–70.
- ^ Vandermeersch, B.; Garralda, M. D. (2011). "Neanderthal geographical and chronological variation". In S. Condemi; G.-C. Weniger (eds.). Continuity and discontinuity in the peopling of Europe. Vertebrate Paleobiology and Paleoanthropology. Springer Netherlands. pp. 113–125. doi:10.1007/978-94-007-0492-3_10. ISBN 978-94-007-0491-6.
- ^ Richter, J. (2011). "When did the Middle Paleolithic begin?". In Conard, N. J.; Richter, J. (eds.). Neanderthal lifeways, subsistence and technology: one hundred and fifty years of Neanderthal study. Vertebrate paleobiology and paleoanthropology. 19. Springer. pp. 7–14. doi:10.1007/978-94-007-0415-2_2. ISBN 978-94-007-0414-5.
- ^ Zanolli, C.; Martinón-Torres, M.; Bernardini, F.; et al. (2018). "The Middle Pleistocene (MIS 12) human dental remains from Fontana Ranuccio (Latium) and Visogliano (Friuli-Venezia Giulia), Italy. A comparative high resolution endostructural assessment". PLOS ONE. 13 (10): e0189773. Bibcode:2018PLoSO..1389773Z. doi:10.1371/journal.pone.0189773. PMC 6169847. PMID 30281595.
- ^ a b c Endicott, P.; Ho, S. Y. W.; Stringer, C. (2010). "Using genetic evidence to evaluate four palaeoanthropological hypotheses for the timing of Neanderthal and modern human origins" (PDF). Journal of Human Evolution. 59 (1): 87–95. doi:10.1016/j.jhevol.2010.04.005. PMID 20510437.
- ^ Welker, F.; et al. (2020). "The dental proteome of Homo antecessor". Nature. 580 (7802): 235–238. Bibcode:2020Natur.580..235W. doi:10.1038/s41586-020-2153-8. PMC 7582224. PMID 32269345. S2CID 214736611.
- ^ Briggs, A. W.; Good, J. M.; Green, R. E. (2009). "Targeted retrieval and analysis of five Neandertal mtDNA genomes" (PDF). Science. 325 (5, 938): 318–321. Bibcode:2009Sci...325..318B. doi:10.1126/science.1174462. PMID 19608918. S2CID 7117454. Archived from the original (PDF) on March 7, 2019.
- ^ Hajdinjak, M.; Fu, Q.; Hübner, A.; et al. (March 1, 2018). "Reconstructing the genetic history of late Neanderthals". Nature. 555 (7698): 652–656. Bibcode:2018Natur.555..652H. doi:10.1038/nature26151. ISSN 1476-4687. PMC 6485383. PMID 29562232.
- ^ a b Krings, M.; Stone, A.; Schmitz, R. W.; Krainitzki, H.; Stoneking, M.; Pääbo, S. (1997). "Neandertal DNA sequences and the origin of modern humans". Cell. 90 (1): 19–30. doi:10.1016/s0092-8674(00)80310-4. hdl:11858/00-001M-0000-0025-0960-8. PMID 9230299. S2CID 13581775.
- ^ Krings, M.; Geisert, H.; Schmitz, R. W.; Krainitzki, H.; Pääbo, S. (1999). "DNA sequence of the mitochondrial hypervariable region II from the Neandertal type specimen". Proceedings of the National Academy of Sciences. 96 (10): 5581–5585. Bibcode:1999PNAS...96.5581K. doi:10.1073/pnas.96.10.5581. PMC 21903. PMID 10318927.
- ^ Green, R. E.; Malaspinas, A. S.; Krause, J.; Briggs, A. W.; et al. (2008). "A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing". Cell. 134 (3): 416–426. doi:10.1016/j.cell.2008.06.021. PMC 2602844. PMID 18692465.
- ^ Sawyer, S.; Renaud, G.; Viola, B.; Hublin, J. J. (2015). "Nuclear and mitochondrial DNA sequences from two Denisovan individuals". Proceedings of the National Academy of Sciences. 112 (51): 15696–15700. Bibcode:2015PNAS..11215696S. doi:10.1073/pnas.1519905112. PMC 4697428. PMID 26630009.
- ^ a b Rogers, A. R.; Harris, N. S.; Achenbach, A. A. (2020). "Neanderthal-Denisovan ancestors interbred with a distantly related hominin". Science Advances. 6 (8): eaay5483. Bibcode:2020SciA....6.5483R. doi:10.1126/sciadv.aay5483. PMC 7032934. PMID 32128408.
- ^ Serangeli, J.; Bolus, M. (2008). "Out of Europe - The dispersal of a successful European hominin form" (PDF). Quartär. 55: 83–98.
- ^ Callander, J. (2004). "Dorothy Garrod's excavations in the Late Mousterian of Shukbah Cave in Palestine reconsidered". Proceedings of the Prehistoric Society. 70: 207–231. doi:10.1017/S0079497X00001171. S2CID 191630165.
- ^ Smith, T. M.; Tafforeau, P.; Reid, D. J.; et al. (2007). "Earliest evidence of modern human life history in North African early Homo sapiens". Proceedings of the National Academy of Sciences of the United States of America. 104 (15): 6128–6133. Bibcode:2007PNAS..104.6128S. doi:10.1073/pnas.0700747104. PMC 1828706. PMID 17372199.
- ^ Douka, K.; J., Zenobia; Lane, C.; et al. (2014). "The chronostratigraphy of the Haua Fteah cave (Cyrenaica, northeast Libya)". Journal of Human Evolution. 66: 39–63. doi:10.1016/j.jhevol.2013.10.001. PMID 24331954.
- ^ Wu, X.-J.; Bruner, E. (2016). "The endocranial anatomy of Maba 1". American Journal of Physical Anthropology. 160 (4): 633–643. doi:10.1002/ajpa.22974. PMID 26972814.
- ^ Nielsen, T. K.; Benito, B. M.; et al. (2017). "Investigating Neanderthal dispersal above 55°N in Europe during the Last Interglacial Complex". Quaternary International. 431: 88–103. Bibcode:2017QuInt.431...88N. doi:10.1016/j.quaint.2015.10.039.
- ^ Nielsen, T. K.; Riede, F. (2018). "On research history and Neanderthal occupation at its northern margins". European Journal of Archaeology. 21 (4): 506–527. doi:10.1017/eaa.2018.12. S2CID 165849999.
- ^ Pavlov, P.; Roebroeks, W.; Svendsen, J. I. (2004). "The Pleistocene colonization of northeastern Europe: a report on recent research". Journal of Human Evolution. 47 (1–2): 3–17. doi:10.1016/j.jhevol.2004.05.002. PMID 15288521.
- ^ a b Slimak, L.; Svendsen, J. I.; Mangerud, J.; Plisson, H. (2011). "Late Mousterian persistence near the Arctic Circle". Science. 332 (6031): 841–845. Bibcode:2011Sci...332..841S. doi:10.1126/science.1203866. JSTOR 29784275. PMID 21566192. S2CID 24688365.
- ^ Slimak, L. (2012). "Response to "Comment on Late Mousterian persistence near the Arctic Circle"". Science. 335 (6065): 167. Bibcode:2012Sci...335..167S. doi:10.1126/science.1210211. PMID 22246757.
- ^ a b Zwyns, N. (2012). "Comment on Late Mousterian persistence near the Arctic Circle". Science. 335 (6065): 167. Bibcode:2012Sci...335..167Z. doi:10.1126/science.1209908. PMID 22246757.
- ^ Holan, S. R.; Deméré, T. A.; Fisher, D. C.; Fullager, R.; Paces, J. B.; Jefferson, G. T. (2017). "A 130,000-year-old archaeological site in southern California, USA". Nature. 544 (7651): 479–483. Bibcode:2017Natur.544..479H. doi:10.1038/nature22065. PMID 28447646.
- ^ Sutton, M. Q.; Parkinson, J.; Rosen, M. D. (2019). "Observations regarding the Cerutti Mastodon". PaleoAmerica. 5 (1): 8–15. doi:10.1080/20555563.2019.1589409. S2CID 155596679.
- ^ Eren, M. I.; Bebber, M. R. (2019). "The Cerutti Mastodon site and experimental archaeology's quiet coming of age". Antiquity. 93 (369): 796–797. doi:10.15184/aqy.2019.50. S2CID 197853460.
- ^ Ferrel, P. M. (2019). "The Cerutti Mastodon site reinterpreted with reference to freeway construction plans and methods". PaleoAmerica. 5 (1): 1–7. doi:10.1080/20555563.2019.1589663. S2CID 167172979.
- ^ a b c d e f g h i Bocquet-Appel, J.; Degioanni, A. (2013). "Neanderthal demographic estimates". Current Anthropology. 54: 202–214. doi:10.1086/673725. S2CID 85090309.
- ^ Hershkovitz, I.; May, H.; Sarig, R.; et al. (2021). "A Middle Pleistocene Homo from Nesher Ramla, Israel". Science. 372 (6549): 1424–1428. Bibcode:2021Sci...372.1424H. doi:10.1126/science.abh3169. S2CID 235628111.
- ^ a b c Mafessoni, F.; Prüfer, K. (2017). "Better support for a small effective population size of Neandertals and a long shared history of Neandertals and Denisovans". Proceedings of the National Academy of Sciences. 114 (48): 10256–10257. doi:10.1073/pnas.1716918114. PMC 5715791. PMID 29138326.
- ^ Lalueza-Fox, C.; Sampietro, M. L.; Caramelli, D.; Puder, Y. (2013). "Neandertal evolutionary genetics: mitochondrial DNA data from the iberian peninsula". Molecular Biology and Evolution. 22 (4): 1077–1081. doi:10.1093/molbev/msi094. PMID 15689531.
- ^ Mellars, P.; French, J. C. (2011). "Tenfold population increase in Western Europe at the Neandertal-to-modern human transition". Science. 333 (6042): 623–627. Bibcode:2011Sci...333..623M. doi:10.1126/science.1206930. PMID 21798948. S2CID 28256970.
- ^ a b Trinkaus, E. (2011). "Late Pleistocene adult mortality patterns and modern human establishment". Proceedings of the National Academy of Sciences. 108 (4): 1267–1271. Bibcode:2011PNAS..108.1267T. doi:10.1073/pnas.1018700108. PMC 3029716. PMID 21220336.
- ^ a b Pettitt, R. B. (2000). "Neanderthal lifecycles: developmental and social phases in the lives of the last archaics". World Archaeology. 31 (3): 351–366. doi:10.1080/00438240009696926. JSTOR 125106. PMID 16475295. S2CID 43859422.
- ^ a b c Gómez-Olivencia, A.; Barash, A.; García-Martínez, D.; et al. (2018). "3D virtual reconstruction of the Kebara 2 Neandertal thorax". Nature Communications. 9 (4387): 4387. Bibcode:2018NatCo...9.4387G. doi:10.1038/s41467-018-06803-z. PMC 6207772. PMID 30377294.
- ^ a b c Trinkaus, E. (1981). "Neanderthal limb proportions and cold adaptation". In Stringer, C. B. (ed.). Aspects of human evolution. Taylor and Francis Ltd.
- ^ Formicola, V.; Giannecchini, M. (1998). "Evolutionary trends of stature in Upper Paleolithic and Mesolithic Europe". Journal of Human Evolution. 36 (3): 325. doi:10.1006/jhev.1998.0270. PMID 10074386.
- ^ Roser, M.; Appel, C.; Ritchie, H. (2013). "Human height". Our World in Data. Retrieved June 16, 2020.
- ^ a b c Duveau, J.; Berillon, G.; Verna, C.; Laisné, G.; Cliquet, D. (2019). "The composition of a Neandertal social group revealed by the hominin footprints at Le Rozel (Normandy, France)". Proceedings of the National Academy of Sciences. 116 (39): 19409–19414. doi:10.1073/pnas.1901789116. PMC 6765299. PMID 31501334.
- ^ Froehle, A. W.; Churchill, S. E. (2009). "Energetic competition between Neandertals and anatomically modern humans" (PDF). PaleoAnthropology: 96–116.
- ^ Gómez-Olivencia, A.; Been, E.; Arsuaga, J. L.; Stock, J. T. (2013). "The Neandertal vertebral column 1: the cervical spine". Journal of Human Evolution. 64 (6): 604–630. doi:10.1016/j.jhevol.2013.02.008. PMID 23541382.
- ^ García-Martínez, D.; Bastir, M.; Huguet, R. (2017). "The costal remains of the El Sidrón Neanderthal site (Asturias, northern Spain) and their importance for understanding Neanderthal thorax morphology". Journal of Human Evolution. 111: 85–101. doi:10.1016/j.jhevol.2017.06.003. hdl:10261/158977. PMID 28874276.
- ^ Gómez-Olivencia, A.; Holliday, T.; Madelaine, M. (2018). "The costal skeleton of the Regourdou 1 Neandertal". Journal of Human Evolution. 130: 151–171. doi:10.1016/j.jhevol.2017.12.005. PMID 29496322.
- ^ a b Weaver, T. D. (2009). "The meaning of Neandertal skeletal morphology". Proceedings of the National Academy of Sciences. 106 (38): 16, 028–16, 033. doi:10.1073/pnas.0903864106. PMC 2752516. PMID 19805258.
- ^ Holliday, T. W. (1997). "Postcranial evidence of cold adaptation in European Neandertals". American Journal of Physical Anthropology. 104 (2): 245–258. doi:10.1002/(SICI)1096-8644(199710)104:2<245::AID-AJPA10>3.0.CO;2-#. PMID 9386830.
- ^ De Groote, I. (2011). "The Neanderthal lower arm". Journal of Human Evolution. 61 (4): 396–410. doi:10.1016/j.jhevol.2011.05.007. PMID 21762953.
- ^ Walker, M. J.; Ortega, J.; Parmová, K.; López, M. V.; Trinkaus, E. (2011). "Morphology, body proportions, and postcranial hypertrophy of a Neandertal female from the Sima de las Palomas, southeastern Spain". Proceedings of the National Academy of Sciences. 108 (25): 10, 087–10, 091. Bibcode:2011PNAS..10810087W. doi:10.1073/pnas.1107318108. PMC 3121844. PMID 21646528.
- ^ Raichlen, D.; Armstrong, H.; Lieberman, D. E. (2011). "Calcaneus length determines running economy: Implications for endurance running performance in modern humans and Neandertals". Journal of Human Evolution. 60 (3): 299–308. doi:10.1016/j.jhevol.2010.11.002. PMID 21269660.
- ^ Lee, S. S. M.; Piazza, S. J. (2009). "Built for speed: musculoskeletal structure and sprinting ability" (PDF). Journal of Experimental Biology. 212 (22): 3700–3707. doi:10.1242/jeb.031096. PMID 19880732. S2CID 10139786. Archived from the original (PDF) on December 14, 2019.
- ^ "Cro-Magnons conquered Europe, but left Neanderthals alone". PLOS Biology. 2 (12): e449. 2004. doi:10.1371/journal.pbio.0020449. PMC 532398.
- ^ Gunz, P.; Tilot, A. K.; Wittfeld, K.; et al. (2019). "Neandertal introgression sheds light on modern human endocranial globularity". Current Biology. 29 (1): 120–127. doi:10.1016/j.cub.2018.10.065. PMC 6380688. PMID 30554901.
- ^ Gunz, P.; Harvati, K. (2007). "The Neanderthal "chignon": variation, integration, and homology". Journal of Human Evolution. 52 (3): 262–274. doi:10.1016/j.jhevol.2006.08.010. PMID 17097133.
- ^ a b c Pearce, E.; Stringer, C.; Dunbar, R. I. M. (2013). "New insights into differences in brain organization between Neanderthals and anatomically modern humans". Proceedings of the Royal Society B. 280 (1758): 20130168. doi:10.1098/rspb.2013.0168. PMC 3619466. PMID 23486442.
- ^ a b c d e Clement, A. F.; Hillson, S. W.; Aiello, L. C. (2012). "Tooth wear, Neanderthal facial morphology and the anterior dental loading hypothesis". Journal of Human Evolution. 62 (3): 367–376. doi:10.1016/j.jhevol.2011.11.014. PMID 22341317.
- ^ Holton, N. E.; Yokley, T. R.; Franciscus, R. G. (2011). "Climatic adaptation and Neandertal facial evolution: A comment on Rae et al. (2011)". Journal of Human Evolution. 61 (5): 624–627. doi:10.1016/j.jhevol.2011.08.001. PMID 21920585.
- ^ O'Connor, C. F.; Franciscus, R. G.; Holton, N. E. (2005). "Bite force production capability and efficiency in Neandertals and modern humans". American Journal of Physical Anthropology. 127 (2): 129–151. doi:10.1002/ajpa.20025. PMID 15558614.
- ^ Benazzi, S.; Nguyen, H. H.; Kullmer, O.; Hublin, J.-J. (2014). "Exploring the biomechanics of taurodontism". Journal of Anatomy. 226 (2): 180–188. doi:10.1111/joa.12260. PMC 4304574. PMID 25407030.
- ^ Takaki, P.; Vieira, M.; Bommarito, S. (2014). "Maximum bite force analysis in different age groups". International Archives of Otorhinolaryngology. 18 (3): 272–276. doi:10.1055/s-0034-1374647. PMC 4297017. PMID 25992105.
- ^ Kleinhubbert, G. (August 10, 2021). "Mister N. bekommt eine dunklere Hautfarbe" [Mister N. gets a darker skin colour]. Der Spiegel (in German). Retrieved August 10, 2021.
- ^ Beals, K.; Smith, C.; Dodd, S. (1984). "Brain size, cranial morphology, climate, and time machines" (PDF). Current Anthropology. 12 (3): 301–30. doi:10.1086/203138. S2CID 86147507.
- ^ Allen, J. S.; Damasio, H.; Grabowski, T. J. (2002). "Normal neuroanatomical variation in the human brain: an MRI-volumetric study". American Journal of Physical Anthropology. 118 (4): 341–358. doi:10.1002/ajpa.10092. PMID 12124914. S2CID 21705705.
- ^ Balzeau, A.; Grimaud-Hervé, D.; Detroit, F.; Holloway, R. L. (2013). "First description of the Cro-Magnon 1 endocast and study of brain variation and evolution in anatomically modern Homo sapiens". Bulletins et Mémoires de la Société d anthropologie de Paris. 25 (1–2): 11–12. doi:10.1007/s13219-012-0069-z. S2CID 14675512.
- ^ Ponce de León, M. S.; Golovanova, L.; Doronichev, V. (2008). "Neanderthal brain size at birth provides insights into the evolution of human life history". Proceedings of the National Academy of Sciences. 105 (37): 13764–13768. doi:10.1073/pnas.0803917105. PMC 2533682. PMID 18779579.
- ^ Kochiyama, T.; Ogihara, N.; Tanabe, H. C. (2018). "Reconstructing the Neanderthal brain using computational anatomy". Scientific Reports. 8 (6296): 6296. Bibcode:2018NatSR...8.6296K. doi:10.1038/s41598-018-24331-0. PMC 5919901. PMID 29700382.
- ^ a b c d e Dannemann, M.; Kelso, J. (2017). "The contribution of Neanderthals to phenotypic variation in modern humans". The American Journal of Human Genetics. 101 (4): 578–589. doi:10.1016/j.ajhg.2017.09.010. PMC 5630192. PMID 28985494.
- ^ a b Allentoft, M. E.; Sikora, M. (2015). "Population genomics of Bronze Age Eurasia". Nature. 522 (7, 555): 167–172. Bibcode:2015Natur.522..167A. doi:10.1038/nature14507. PMID 26062507. S2CID 4399103.
- ^ Cerqueira, C. C.; Piaxão-Côrtes, V. R.; Zambra, F. M. B.; Hünemeier, T.; Bortolini, M. (2012). "Predicting Homo pigmentation phenotype through genomic data: From neanderthal to James Watson". American Journal of Human Biology. 24 (5): 705–709. doi:10.1002/ajhb.22263. PMID 22411106. S2CID 25853632.
- ^ Lalueza-Fox, C.; Rompler, H.; Caramelli, D.; et al. (2007). "A melanocortin 1 receptor allele suggests varying pigmentation among Neanderthals". Science. 318 (5855): 1453–1455. Bibcode:2007Sci...318.1453L. doi:10.1126/science.1147417. PMID 17962522. S2CID 10087710.
- ^ Smith, T. M.; Tafforeau, P.; Reid, D. J. (2010). "Dental evidence for ontogenetic differences between modern humans and Neanderthals". Proceedings of the National Academy of Sciences. 107 (49): 20923–20928. Bibcode:2010PNAS..10720923S. doi:10.1073/pnas.1010906107. PMC 3000267. PMID 21078988.
- ^ Guatelli-Steinberg, D. (2009). "Recent studies of dental development in Neandertals: Implications for Neandertal life histories". Evolutionary Biology. 18 (1): 9–20. doi:10.1002/evan.20190. S2CID 84273699.
- ^ Rosas, A.; Ríos, L.; Estalrrich, A.; et al. (2017). "The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain)". Science. 357 (6537): 1282–1287. Bibcode:2017Sci...357.1282R. doi:10.1126/science.aan6463. PMID 28935804.
- ^ DeSilva, J. M. (2018). "Comment on 'The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain)'". Science. 359 (6380): eaar3611. doi:10.1126/science.aar3611. PMID 29590012.
- ^ Froehle, A. W.; Churchill, S. E. (2009). "Energetic competition between Neanderthals and anatomically modern humans" (PDF). Paleoanthropology: 96–116.
- ^ a b Snodgrass, J. J.; Leonard, W. R. (2009). "Neandertal energetics revisited: insights into population dynamics and life history evolution" (PDF). Paleoanthropology: 220–237. doi:10.4207/PA.2009.ART31. S2CID 16044193. Archived from the original (PDF) on August 1, 2020.
- ^ a b c Hockett, B. (2012). "The consequences of Middle Paleolithic diets on pregnant Neanderthal women". Quaternary International. 264: 78–82. Bibcode:2012QuInt.264...78H. doi:10.1016/j.quaint.2011.07.002.
- ^ Taylor, J. S.; Reimchen, T. E. (2016). "Opsin gene repertoires in northern archaic hominids". Genome. 59 (8): 541–549. doi:10.1139/gen-2015-0164. hdl:1807/73317. PMID 27463216.
- ^ a b Nakahashi, W. (2017). "The effect of trauma on Neanderthal culture: A mathematical analysis". Homo. 68 (2): 83–100. doi:10.1016/j.jchb.2017.02.001. PMID 28238406.
- ^ Trinkaus, E.; Villotte, S (2017). "External auditory exostoses and hearing loss in the Shanidar 1 Neandertal". PLOS ONE. 12 (10): e0186684. Bibcode:2017PLoSO..1286684T. doi:10.1371/journal.pone.0186684. PMC 5650169. PMID 29053746.
- ^ Trinkaus, E.; Samsel, M.; Villotte, S. (2019). "External auditory exostoses among western Eurasian late Middle and Late Pleistocene humans". PLOS ONE. 14 (8): e0220464. Bibcode:2019PLoSO..1420464T. doi:10.1371/journal.pone.0220464. PMC 6693685. PMID 31412053.
- ^ Beier, J.; Anthes, N.; Wahl, J.; Harvati, K. (2018). "Similar cranial trauma prevalence among Neanderthals and Upper Palaeolithic modern humans". Nature. 563 (7733): 686–690. Bibcode:2018Natur.563..686B. doi:10.1038/s41586-018-0696-8. PMID 30429606. S2CID 53306963.
- ^ Trinkaus, E. (2012). "Neandertals, early modern humans, and rodeo riders". Journal of Archaeology. 39 (2): 3691–3693. doi:10.1016/j.jas.2012.05.039.
- ^ a b Camarós, E.; Cueto, M.; Lorenzo, C.; Villaverde, V. (2016). "Large carnivore attacks on hominins during the Pleistocene: a forensic approach with a Neanderthal example". Archaeological and Anthropological Sciences. 8 (3): 635–646. doi:10.1007/s12520-015-0248-1. hdl:10550/54275. S2CID 82001651.
- ^ Sánchez-Quinto, F.; Lalueza-Fox, C. (2015). "Almost 20 years of Neanderthal palaeogenetics: adaptation, admixture, diversity, demography and extinction". Philosophical Transactions of the Royal Society B. 370 (1660): 20130374. doi:10.1098/rstb.2013.0374. PMC 4275882. PMID 25487326.
- ^ a b c Ríos, L.; Kivell, T. L.; Lalueza-Fox, C.; Estalrrich, A. (2019). "Skeletal anomalies in the Neandertal family of El Sidrón (Spain) support a role of inbreeding in Neandertal extinction". Scientific Reports. 9 (1): 1697. Bibcode:2019NatSR...9.1697R. doi:10.1038/s41598-019-38571-1. PMC 6368597. PMID 30737446.
- ^ Haeusler, M.; Trinkaus, E.; Fornai, C.; Müller, J.; Bonneau, N.; Boeni, T.; Frater, N. (2019). "Morphology, pathology, and the vertebral posture of the La Chapelle-aux-Saints Neandertal". Proceedings of the National Academy of Sciences. 116 (11): 4923–4927. doi:10.1073/pnas.1820745116. PMC 6421410. PMID 30804177.
- ^ Crubézy, E.; Trinkaus, E. (1992). "Shanidar 1: a case of hyperostotic disease (DISH) in the middle Paleolithic". American Journal of Physical Anthropology. 89 (4): 411–420. doi:10.1002/ajpa.1330890402. PMID 1463085.
- ^ Houldcroft, C. J.; Underdown, S. J. (2016). "Neanderthal genomics suggests a pleistocene time frame for the first epidemiologic transition" (PDF). American Journal of Physical Anthropology. 160 (3): 379–388. doi:10.1002/ajpa.22985. PMID 27063929.
- ^ Pimenoff, V. N.; de Oliveira, C. M.; Bravo, I. G. (2017). "Transmission between archaic and modern human ancestors during the evolution of the oncogenic human papillomavirus 16". Molecular Biology and Evolution. 34 (1): 4–19. doi:10.1093/molbev/msw214. PMC 5854117. PMID 28025273.
- ^ a b c d Weyrich, L. S.; Duchene, S.; Soubrier, J.; et al. (2017). "Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus". Nature. 544 (7650): 357–361. Bibcode:2017Natur.544..357W. doi:10.1038/nature21674. hdl:10261/152016. PMID 28273061. S2CID 4457717.
- ^ Fennel, K. J.; Trinkaus, E. (1997). "Bilateral femoral and tibial periostitis in the La Ferrassie 1 Neanderthal". Journal of Archaeological Science. 24 (11): 985–995. doi:10.1006/jasc.1996.0176.
- ^ Soltysiak, A. (2012). "Comment: low dental caries rate in Neandertals: the result of diet or the oral flora composition?". Homo. 63 (2): 110–113. doi:10.1016/j.jchb.2012.02.001. PMID 22409830.
- ^ a b c Smith, T. A.; Austin, C.; Green, D. R.; et al. (2018). "Wintertime stress, nursing, and lead exposure in Neanderthal children". Science Advances. 4 (10): eaau9483. Bibcode:2018SciA....4.9483S. doi:10.1126/sciadv.aau9483. PMC 6209393. PMID 30402544.
- ^ Rosas, A.; Estalrrich, A.; García-Vargas, S.; García-Tabernero, A.; Huguet, R.; Lalueza-Fox, C.; de la Rasilla, M. (2013). "Identification of Neandertal individuals in fragmentary fossil assemblages by means of tooth associations: The case of El Sidrón (Asturias, Spain)". Comptes Rendus Palevol. 12 (5): 279–291. doi:10.1016/j.crpv.2013.06.003.
- ^ a b c d e Spikins, P.; Hitchens, G.; Needham, A.; et al. (2014). "The Cradle of Thought: Growth, Learning, Play and Attachment in Neanderthal Children" (PDF). Oxford Journal of Archaeology. 33 (2): 111–134. doi:10.1111/ojoa.12030.
- ^ Farizy, C. (1994). "Spatial patterning of Middle Paleolithic sites". Journal of Anthropological Archaeology. 13 (2): 153–160. doi:10.1006/jaar.1994.1010.
- ^ Fortes, G. G.; Grandal-d'Anglade, A.; Kolbe, B. (2016). "Ancient DNA reveals differences in behaviour and sociality between brown bears and extinct cave bears" (PDF). Molecular Ecology. 25 (19): 4907–4918. doi:10.1111/mec.13800. PMID 27506329. S2CID 18353913.
- ^ Stiller, M.; Baryshnikov, G.; Bocherens, H. (2010). "Withering away—25,000 years of genetic decline preceded cave bear extinction". Molecular Biology and Evolution. 27 (5): 975–978. doi:10.1093/molbev/msq083. PMID 20335279.
- ^ Ekshtain, R.; Malinsky-Buller, A.; Greenbaum, N.; et al. (2019). "Persistent Neanderthal occupation of the open-air site of 'Ein Qashish, Israel". PLOS ONE. 14 (6): e0215668. Bibcode:2019PLoSO..1415668E. doi:10.1371/journal.pone.0215668. PMC 6594589. PMID 31242180.
- ^ Been, E.; Hovers, E.; Ekshtain, R. (2017). "The first Neanderthal remains from an open-air Middle Palaeolithic site in the Levant". Scientific Reports. 7 (2958): 2958. Bibcode:2017NatSR...7.2958B. doi:10.1038/s41598-017-03025-z. PMC 5462778. PMID 28592838.
- ^ Lalueza-Fox, C.; Rosas, A.; Estalrrich, A. (2011). "Genetic evidence for patrilocal mating behavior among Neandertal groups". Proceedings of the National Academy of Sciences. 108 (1): 250–253. doi:10.1073/pnas.1011553108. PMC 3017130. PMID 21173265.
- ^ Pearce, E.; Moutsiou, T. (2015). "Using obsidian transfer distances to explore social network maintenance in late Pleistocene hunter-gatherers". Journal of Anthropology and Archaeology. 36: 12–20. doi:10.1016/j.jaa.2014.07.002. PMC 4157217. PMID 25214705.
- ^ a b Rosas, A.; Bastir, M.; Martínez-Maza, C. (2006). "Paleobiology and comparative morphology of a late Neandertal sample from El Sidrón, Asturias, Spain". Proceedings of the National Academy of Sciences. 103 (51): 19266–19271. Bibcode:2006PNAS..10319266R. doi:10.1073/pnas.0609662104. PMC 1748215. PMID 17164326.
- ^ a b Kolobova, K. A.; Roberts, R. G.; Chabai, V. P.; et al. (2020). "Archaeological evidence for two separate dispersals of Neanderthals into southern Siberia". PNAS. 117 (6): 2979–2885. doi:10.1073/pnas.1918047117. PMC 7022189. PMID 31988114.
- ^ Picin, A.; Hajdinjak, M.; Nowaczewska, W. (2020). "New perspectives on Neanderthal dispersal and turnover from Stajnia Cave (Poland)". Scientific Reports. 10 (14778): 14778. Bibcode:2020NatSR..1014778P. doi:10.1038/s41598-020-71504-x. PMC 7479612. PMID 32901061.
- ^ Zollikofer, C. P. E.; Ponce de Leon, M. S.; Vandermeersch, B.; Leveque, F. (2002). "Evidence for interpersonal violence in the St. Cesaire Neanderthal". Proceedings of the National Academy of Sciences. 99 (9): 6444–6448. Bibcode:2002PNAS...99.6444Z. doi:10.1073/pnas.082111899. PMC 122968. PMID 11972028.
- ^ Churchill, S. E.; Franciscus, R. G.; McKean-Peraza, H. A.; Daniel, J. A.; Warren, B. R. (2009). "Shanidar 3 Neandertal rib puncture wound and Paleolithic weaponry". Journal of Human Evolution. 57 (2): 163–178. doi:10.1016/j.jhevol.2009.05.010. PMID 19615713.
- ^ Estalrrich, A.; Rosas, A. (2015). "Division of labor by sex and age in Neandertals: an approach through the study of activity-related dental wear". Journal of Human Evolution. 80: 51–63. doi:10.1016/j.jhevol.2014.07.007. PMID 25681013.
- ^ a b c Camarós, E.; Cueto, M.; Teira, L.; Münzel, S. C. (2017). "Bears in the scene: Pleistocene complex interactions with implications concerning the study of Neanderthal behavior". Quaternary International. 435: 237–246. Bibcode:2017QuInt.435..237C. doi:10.1016/j.quaint.2015.11.027.
- ^ Bocherens, H.; Drucker, D. G.; Billiou, D.; Patou-Mathis, M.; Vandermeersch, B. (2005). "Isotopic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal: review and use of a multi-source mixing model". Journal of Human Evolution. 49 (1): 71–87. doi:10.1016/j.jhevol.2005.03.003. PMID 15869783.
- ^ Jaouen, K.; et al. (2019). "Exceptionally high δ15N values in collagen single amino acids confirm Neandertals as high-trophic level carnivores". Proceedings of the National Academy of Sciences. 116 (11): 4928–4933. doi:10.1073/pnas.1814087116. PMC 6421459. PMID 30782806.
- ^ Scott, K. (1980). "Two hunting episodes of middle Palaeolithic age at La Cotte de Saint‐Brelade, Jersey (Channel Islands)". World Archaeology. 12 (2): 137–152. doi:10.1080/00438243.1980.9979788.
- ^ a b Scott, B.; Bates, M.; Bates, C. R.; Conneller, C. (2015). "A new view from La Cotte de St Brelade, Jersey". Antiquity. 88 (339): 13–29. doi:10.1017/S0003598X00050195.
- ^ a b Marín, J.; Saladié, P.; Rodríguez-Hidalgo, A.; Carbonell, E. (2017). "Neanderthal hunting strategies inferred from mortality profiles within the Abric Romaní sequence". PLOS ONE. 12 (11): e0186970. Bibcode:2017PLoSO..1286970M. doi:10.1371/journal.pone.0186970. PMC 5699840. PMID 29166384.
- ^ a b El Zaatari, S.; Grine, F. E.; Ungar, P. S.; Hublin, J.-J. (2016). "Neandertal versus modern human dietary responses to climatic fluctuations". PLOS ONE. 11 (4): e0153277. Bibcode:2016PLoSO..1153277E. doi:10.1371/journal.pone.0153277. PMC 4847867. PMID 27119336.
- ^ Hardy, B. L. (2010). "Climatic variability and plant food distribution in Pleistocene Europe: Implications for Neanderthal diet and subsistence". Quaternary Science Reviews. 29 (5–6): 662–679. Bibcode:2010QSRv...29..662H. doi:10.1016/j.quascirev.2009.11.016.
- ^ Buck, L. T.; Stringer, C. B. (2014). "Having the stomach for it: a contribution to Neanderthal diets?" (PDF). Quaternary Science Reviews. 96: 161–167. Bibcode:2014QSRv...96..161B. doi:10.1016/j.quascirev.2013.09.003.
- ^ Fiorenza, L.; Benazzi, S.; Tausch, J.; Kullmer, O.; Bromage, T. G.; Schrenk, F. (2011). Rosenberg, Karen (ed.). "Molar macrowear reveals Neanderthal eco-geographic dietary variation". PLOS ONE. 6 (3): e14769. Bibcode:2011PLoSO...614769F. doi:10.1371/journal.pone.0014769. PMC 3060801. PMID 21445243.
- ^ Romandini, M.; Terlato, G.; Nannini, N. (2018). "Bears and humans, a Neanderthal tale. Reconstructing uncommon behaviors from zooarchaeological evidence in southern Europe". Journal of Archaeological Science. 90: 71–91. doi:10.1016/j.jas.2017.12.004. hdl:11392/2381729.
- ^ Colonese, A. C.; Mannino, M. A.; Mayer, DE (2011). "Marine mollusc exploitation in Mediterranean prehistory: an overview". Quaternary International. 239 (1–2): 86–103. Bibcode:2011QuInt.239...86C. doi:10.1016/j.quaint.2010.09.001.
- ^ a b c Villa, P.; Soriano, S.; Pollarolo, L. (2020). "Neandertals on the beach: use of marine resources at Grotta dei Moscerini (Latium, Italy)". PLOS ONE. 15 (1): e0226690. Bibcode:2020PLoSO..1526690V. doi:10.1371/journal.pone.0226690. PMC 6961883. PMID 31940356.
- ^ Cortés-Sánchez, M.; Morales-Muñiz, A.; et al. (2011). "Earliest known use of marine resources by Neanderthals". PLOS ONE. 6 (9): e24026. Bibcode:2011PLoSO...624026C. doi:10.1371/journal.pone.0024026. PMC 3173367. PMID 21935371.
- ^ Stringer, C. B.; Finlayson, C.; Barton, R. N. E. (2008). "Neanderthal exploitation of marine mammals in Gibraltar". Proceedings of the National Academy of Sciences. 105 (38): 14319–14324. Bibcode:2008PNAS..10514319S. doi:10.1073/pnas.0805474105. PMC 2567146. PMID 18809913.
- ^ Zilhão, J.; Angelucci, D. E.; Igreja, M. A.; et al. (2020). "Last Interglacial Iberian Neandertals as fisher-hunter-gatherers" (PDF). Science. 367 (6485): eaaz7943. doi:10.1126/science.aaz7943. PMID 32217702. S2CID 214671143.
- ^ a b c d Hardy, B. L.; Moncel, M.-H.; Daujeard, C.; et al. (2013). "Impossible Neanderthals? Making string, throwing projectiles and catching small game during Marine Isotope Stage 4 (Abri du Maras, France)". Quaternary Science Reviews. 82: 23–40. Bibcode:2013QSRv...82...23H. doi:10.1016/j.quascirev.2013.09.028.
- ^ Bocherens, H.; Baryshnikov, G.; Van Neer, W. (2014). "Were bears or lions involved in salmon accumulation in the Middle Palaeolithic of the Caucasus? An isotopic investigation in Kudaro 3 cave". Quaternary International. 339–340: 112–118. Bibcode:2014QuInt.339..112B. doi:10.1016/j.quaint.2013.06.026.
- ^ Trinkaus, E. (1975). "Squatting among the Neandertals: A problem in the behavioral interpretation of skeletal morphology". Journal of Archaeological Science. 2 (4): 327–351. doi:10.1016/0305-4403(75)90005-9.
- ^ Sistiaga, A.; et al. (2014). "The Neanderthal meal: A new perspective using faecal biomarkers". PLOS ONE. 9 (6): e101045. Bibcode:2014PLoSO...9j1045S. doi:10.1371/journal.pone.0101045. PMC 4071062. PMID 24963925.
- ^ Daugeard, C. (2008). "Exploitation du milieu animal par les Néandertaliens dans le Sud-Est de la France" [Exploitation of the environment by Neanderthals in the southeast of France]. Bulletin de la Société préhistorique française (in French). 106 (4): 818–819.
- ^ a b Speth, J. D. (2015). "When did humans learn to boil?" (PDF). PaleoAnthropology: 54–67.
- ^ Agam, A.; Barkai, R. (2018). "Elephant and mammoth hunting during the Paleolithic: a review of the relevant archaeological, ethnographic and ethnohistorical records". Quaternary. 1 (3): 19. doi:10.3390/quat1010003.
- ^ a b c Zilhão, J.; Angelucci, D. E.; Badal-García, E. (2010). "Symbolic use of marine shells and mineral pigments by Iberian Neandertals". Proceedings of the National Academy of Sciences. 107 (3): 1023–1028. Bibcode:2010PNAS..107.1023Z. doi:10.1073/pnas.0914088107. PMC 2824307. PMID 20080653.
- ^ Diedrich, C. G. (2010). "The Crocuta crocuta spelaea (Goldfuss 1823) population and its prey from the Late Pleistocene Teufelskammer Cave hyena den besides the famous Paleolithic Neandertal Cave (NRW, NW Germany)". Historical Biology. 23 (2–3): 237–270. doi:10.1080/08912963.2010.530348. S2CID 128758869.
- ^ a b c Rougier, H.; Crevecoeur, I.; Beauval, C. (2016). "Neandertal cannibalism and Neandertal bones used as tools in Northern Europe". Scientific Reports. 6 (29, 005): 29005. Bibcode:2016NatSR...629005R. doi:10.1038/srep29005. PMC 4933918. PMID 27381450.
- ^ a b c d Yravedra, J.; Yustos, M. (2015). "Cannibalism in the Neanderthal world: an exhaustive revision". Journal of Taphonomy. 13 (1): 33–52.
- ^ Defleur, A.; White, T.; Valensi, P.; Slimak, L. (1999). "Neanderthal cannibalism at Moula-Guercy, Ardèche, France". Science. 286 (5437): 128–131. doi:10.1126/science.286.5437.128. PMID 10506562.
- ^ Maureille, B.; Sandrine, C.; Beauval, C.; Mann, A. E. (2017). "The challenges of identifying partially digested human teeth: first description of Neandertal remains from the Mousterian site of Marillac (Marillac-le-Franc, Charente, France) and implications for palaeoanthropological research". Paleo. 28: 201–212. doi:10.4000/paleo.3448.
- ^ a b Roebroeks, W.; Sier, M. J.; Nielsen, T. K.; et al. (2012). "Use of red ochre by early Neandertals". Proceedings of the National Academy of Sciences. 109 (6): 1889–1894. Bibcode:2012PNAS..109.1889R. doi:10.1073/pnas.1112261109. PMC 3277516. PMID 22308348.
- ^ d'Errico, F.; Vanhaeren, M.; Henshilwood, C. (2009). "From the origin of language to the diversification of languages: What archaeology and paleoanthropology say?". Becoming eloquent: advances in the emergence of language, human cognition, and modern cultures. John Benjamins Publishing Company. p. 25. ISBN 978-90-272-3269-4.
- ^ Marin, C.; Moncele, M.-H.; Anghelinu, M.; Cârciumaru, R. (2002). "The Cioarei-Boroșteni Cave (Carpathian Mountains, Romania): Middle Palaeolithic finds and technological analysis of the lithic assemblages". Antiquity. 76 (293): 681–690. doi:10.1017/S0003598X00091122.
- ^ Peresani, M.; Vanhaeren, M.; Quaggiotto, E.; et al. (2013). "An ochered fossil marine shell from the Mousterian of Fumane Cave, Italy". PLOS ONE. 8 (7): e68572. Bibcode:2013PLoSO...868572P. doi:10.1371/journal.pone.0068572. PMC 3707824. PMID 23874677.
- ^ Higham, T.; Jacobi, R.; Julien, M.; David, F.; Basell, L.; Wood, R.; Davies, W.; Ramsey, C. B. (2010). "Chronology of the Grotte du Renne (France) and implications for the context of ornaments and human remains within the Chatelperronian". Proceedings of the National Academy of Sciences. 107 (47): 20234–20239. Bibcode:2010PNAS..10720234H. doi:10.1073/pnas.1007963107. PMC 2996711. PMID 20956292.
- ^ Mellars, P. (2010). "Neanderthal symbolism and ornament manufacture: The bursting of a bubble?". Proceedings of the National Academy of Sciences. 107 (47): 20147–20148. Bibcode:2010PNAS..10720147M. doi:10.1073/pnas.1014588107. PMC 2996706. PMID 21078972.
- ^ Hublin, J.-J.; Talamo, S.; Julien, M.; David, F.; Connet, N.; Bodu, P.; Vandermeersch, B.; Richards, M.P. (2012). "Radiocarbon dates from the Grotte du Renne and Saint-Césaire support a Neandertal origin for the Châtelperronian". Proceedings of the National Academy of Sciences USA. 109 (46): 18743–18748. Bibcode:2012PNAS..10918743H. doi:10.1073/pnas.1212924109. PMC 3503158. PMID 23112183.
- ^ Welkera, F.; Hajdinjak, M.; Talamo, S.; Jaouen, K.; Dannemann, M.; David, F.; Julien, M.; Meyer, M.; Kelso, J.; Barnes, I.; Brace, S.; Kamminga, P.; Fischer, R.; Kessler, B.M.; Stewart, J.R.; Pääbo, S.; Collins, M.J.; Hublin, J.-J. (2016). "Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne". Proceedings of the National Academy of Sciences USA. 113 (40): 11162–11167. doi:10.1073/pnas.1605834113. PMC 5056053. PMID 27638212.
- ^ Finlayson 2019, pp. 129–132.
- ^ Finlayson, C.; Brown, K.; Blasco, R.; Rosell, J.; Negro, J. José; Bortolotti, G. R.; Finlayson, G.; Sánchez Marco, A.; Giles Pacheco, F.; Rodríguez Vidal, J.; Carrión, J. S.; Fa, D. A.; Rodríguez Llanes, J. M. (2012). "Birds of a feather: Neanderthal exploitation of raptors and corvids". PLOS ONE. 7 (9): e45927. Bibcode:2012PLoSO...745927F. doi:10.1371/journal.pone.0045927. PMC 3444460. PMID 23029321.
- ^ a b Finlayson, S.; Finlayson, G.; Guzman, F. G.; Finlayson, C. (2019). "Neanderthals and the cult of the sun bird". Quaternary Science Reviews. 217: 217–224. Bibcode:2019QSRv..217..217F. doi:10.1016/j.quascirev.2019.04.010. S2CID 149949579.
- ^ Radovčić, D.; Sršen, A. O.; Radovčić, J.; Frayer, D. W.; Petraglia, M. D. (2015). "Evidence for Neandertal jewelry: modified white-tailed eagle claws at Krapina". PLOS ONE. 10 (3): e0119802. Bibcode:2015PLoSO..1019802R. doi:10.1371/journal.pone.0119802. PMC 4356571. PMID 25760648.
- ^ Callaway, E. (2015). "Neanderthals wore eagle talons as jewellery". Nature. doi:10.1038/nature.2015.17095. S2CID 191368693.
- ^ Rodríguez-Hidalgo, A.; Morales, J. I.; Cebrià, A.; et al. (2019). "The Châtelperronian Neanderthals of Cova Foradada (Calafell, Spain) used imperial eagle phalanges for symbolic purposes". Science Advances. 5 (11): eaax1984. Bibcode:2019SciA....5.1984R. doi:10.1126/sciadv.aax1984. PMC 6824858. PMID 31701003.
- ^ Marquet, J.; Lorblanchet, M.; Oberlin, C.; Thamo-Bozso, E.; Aubry, T. (2016). "New dating of the "mask" of La Roche-Cotard (Langeais, Indre-et-Loire, France)". Paleo Revue d'Archéologie Préhistorique. 27: 253–263.
- ^ Marquet, J.-C.; Lorblanchet, M. (2003). "A Neanderthal face? The proto-figurine from La Roche-Cotard, Langeais (Indre-et-Loire, France)". Antiquity. 77 (298): 661–670. doi:10.1017/s0003598x00061627. ISSN 0003-598X. S2CID 162609308.
- ^ Pettitt, P. B. (2003). "Is this the infancy of art? Or the art of an infant? A possible Neanderthal face from La Roche-Cotard, France" (PDF). Before Farming. 11 (3). Archived from the original (PDF) on October 2, 2011.
- ^ "But is it art?". Science. 302 (5652): 1890a–1890. 2003. doi:10.1126/science.302.5652.1890a. S2CID 149146480.
- ^ Marshack, A. (1988). "The Neanderthals and the human capacity for symbolic thought". In Otte, M. (ed.). L'Homme de Néandertal: la pensée [Neanderthal man: thought]. 5. Université de Liège. p. 71.
- ^ E. Callaway (2014). "Neanderthals made some of Europe's oldest art". Nature News. doi:10.1038/nature.2014.15805. S2CID 163840392.
- ^ Rodríguez-Vidal, J.; d'Errico, F.; Pacheco, F. G. (2014). "A rock engraving made by Neanderthals in Gibraltar". Proceedings of the National Academy of Sciences. 111 (37): 13301–13306. Bibcode:2014PNAS..11113301R. doi:10.1073/pnas.1411529111. PMC 4169962. PMID 25197076.
- ^ Leder, D.; Hermann, R.; Hüls, M.; et al. (2021). "A 51,000-year-old engraved bone reveals Neanderthals' capacity for symbolic behaviour". Nature and Ecology. 5 (9): 1273–1282. doi:10.1038/s41559-021-01487-z. PMID 34226702. S2CID 235746596.
- ^ Marris, E. (2018). "Neanderthal artists made oldest-known cave paintings". Nature News. doi:10.1038/d41586-018-02357-8.
- ^ Cârciumaru, M.; Niţu, E.-C.; Cîrstina, O. (2015). "A geode painted with ochre by the Neanderthal man". Comptes Rendus Palevol. 14 (2): 31–41. doi:10.1016/j.crpv.2014.05.003.
- ^ Dobosi, V. T. (1985). "Jewelry, musical instruments and exotic objects from the Hungarian Palaeolithic". Folia Archaeologica. 36: 7–29.
- ^ Withalm, G. (2004). "New evidence for cave bear hunting from Potočka Zijalka Cave, Slovenia". Mitteilungen der Kommission für Quartärforschung der Österreichischen Akademie der Wissenschaften. 13: 219–234.
- ^ a b Diedrich, C. G. (2015). "'Neanderthal bone flutes': simply products of Ice Age spotted hyena scavenging activities on cave bear cubs in European cave bear dens". Royal Society Open Science. 2 (4): 140022. Bibcode:2015RSOS....240022D. doi:10.1098/rsos.140022. PMC 4448875. PMID 26064624.
- ^ Fink, R. (1997). "Neanderthal flute: oldest musical instrument: matches notes of do, re, mi scale: musicological analysis". Crosscurrents (183): 1–9. ISBN 978-0-912424-12-5.
- ^ Brodar, Mitja (September 26, 2008). ""Piščalka" iz Divjih bab ni neandertalska" [The Divje Babe "Flute" is not Neanderthal] (in Slovenian). Archived from the original on July 28, 2011.
- ^ Richter, D.; Grün, R.; Joannes-Boyau, R.; Steele; et al. (2017). "The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age". Nature. 546 (7657): 293–296. Bibcode:2017Natur.546..293R. doi:10.1038/nature22335. PMID 28593967. S2CID 205255853.
- ^ Li, F.; Kuhn, S. L.; Chen, F. (2018). "The easternmost Middle Paleolithic (Mousterian) from Jinsitai Cave, North China". Journal of Human Evolution. 114: 76–84. doi:10.1016/j.jhevol.2017.10.004. PMID 29447762.
- ^ a b Rolland, N.; Dibble, H. L. (1990). "A new synthesis of Middle Paleolithic variability". American Antiquity. 55 (3): 480–499. doi:10.2307/281279. JSTOR 281279.
- ^ Rocca, R.; Connet, N.; Lhomme, V. (2017). "Before the transition? The final middle Palaeolithic lithic industry from the Grotte du Renne (layer XI) at Arcy-sur-Cure (Burgundy, France)". Comptes Rendus Palevol. 16 (8): 878–893. doi:10.1016/j.crpv.2017.04.003.
- ^ a b Milks, A.; Parker, D.; Pope, M. (2019). "External ballistics of Pleistocene hand-thrown spears: experimental performance data and implications for human evolution". Scientific Reports. 9 (1): 820. Bibcode:2019NatSR...9..820M. doi:10.1038/s41598-018-37904-w. PMC 6347593. PMID 30683877.
- ^ a b Faivre, J.-P.; Maureille, B.; Bayle, P.; Crevecoeur, I. (2014). "Middle Pleistocene human remains from Tourville-la-Rivière (Normandy, France) and their archaeological context". PLOS ONE. 9 (10): e104111. Bibcode:2014PLoSO...9j4111F. doi:10.1371/journal.pone.0104111. PMC 4189787. PMID 25295956.
- ^ Boëda, E.; Geneste, J. M.; Griggo, C.; Mercier, N.; Muhesen, S.; Reyss, J. L.; Taha, A.; Valladas, H. (2015). "A Levallois point embedded in the vertebra of a wild ass (Equus africanus): hafting, projectiles and Mousterian hunting weapons". Antiquity. 73 (280): 394–402. CiteSeerX 10.1.1.453.29. doi:10.1017/S0003598X00088335.
- ^ Roussel, M.; Soressi, M.; Hublin, J.-J. (2016). "The Châtelperronian conundrum: blade and bladelet lithic technologies from Quinçay, France". Journal of Human Evolution. 95: 13–32. doi:10.1016/j.jhevol.2016.02.003. PMID 27260172.
- ^ Hublin, J.-J.; Talamo, S.; Julien, M.; David, F.; Connet, N.; Bodu, P.; Vandermeersch, B.; Richards, M. P. (2012). "Radiocarbon dates from the Grotte du Renne and Saint-Césaire support a Neandertal origin for the Châtelperronian". Proceedings of the National Academy of Sciences. 190 (46): 18743–18748. Bibcode:2012PNAS..10918743H. doi:10.1073/pnas.1212924109. PMC 3503158. PMID 23112183.
- ^ a b Soressi, M.; McPherron, S. P.; Lenoir, M.; et al. (2013). "Neandertals made the first specialized bone tools in Europe". Proceedings of the National Academy of Sciences. 110 (35): 14186–14190. Bibcode:2013PNAS..11014186S. doi:10.1073/pnas.1302730110. PMC 3761603. PMID 23940333.
- ^ Caron, F.; d'Errico, F.; Del Moral, P.; Santos, F.; Zilhâo, J. (2011). "The reality of Neandertal symbolic behavior at the Grotte du Renne, Arcy-sur-Cure, France". PLOS ONE. 6 (6): e21545. Bibcode:2011PLoSO...621545C. doi:10.1371/journal.pone.0021545. PMC 3126825. PMID 21738702.
- ^ Mellars, P.; Gravina, B.; Ramsey, C. B. (2007). "Confirmation of Neanderthal/modern human interstratification at the Chatelperronian type-site". Proceedings of the National Academy of Sciences. 104 (9): 3657–3662. Bibcode:2007PNAS..104.3657M. doi:10.1073/pnas.0608053104. PMC 1805566. PMID 17360698.
- ^ Gravina, B.; Bachellerie, F.; Caux, S. (2018). "No reliable evidence for a Neanderthal-Châtelperronian association at La Roche-à-Pierrot, Saint-Césaire". Scientific Reports. 8 (1): 15134. Bibcode:2018NatSR...815134G. doi:10.1038/s41598-018-33084-9. PMC 6181958. PMID 30310091.
- ^ Villa, P.; Pollarolo, L.; Conforti, J.; et al. (2018). "From Neandertals to modern humans: new data on the Uluzzian". PLOS ONE. 13 (5): e0196786. Bibcode:2018PLoSO..1396786V. doi:10.1371/journal.pone.0196786. PMC 5942857. PMID 29742147.
- ^ a b Hoffecker, J. F. (2009). "The spread of modern humans in Europe". Proceedings of the National Academy of Sciences. 106 (38): 16040–16045. Bibcode:2009PNAS..10616040H. doi:10.1073/pnas.0903446106. PMC 2752585. PMID 19571003.
- ^ Martisius, N. L.; Welker, F.; Dogandžić, T.; et al. (2020). "Non-destructive ZooMS identification reveals strategic bone tool raw material selection by Neandertals". Scientific Reports. 10 (1): 7746. Bibcode:2020NatSR..10.7746M. doi:10.1038/s41598-020-64358-w. PMC 7210944. PMID 32385291.
- ^ Sykes 2020, pp. 260–261.
- ^ Speth, J. D.; Tchernov, E. (2002). "Middle Paleolithic tortoise use at Kebara Cave (Israel)". Journal of Archaeological Science. 29 (5): 471–483. doi:10.1006/jasc.2001.0740.
- ^ Aranguren, B.; Revedin, A.; Amico, N.; Cavulli, F.; Giachi, G.; Grimaldi, S.; Macchioni, N.; Santaniello, F. (2018). "Wooden tools and fire technology in the early Neanderthal site of Poggetti Vecchi (Italy)". Proceedings of the National Academy of Sciences. 115 (9): 2054–2059. doi:10.1073/pnas.1716068115. PMC 5834685. PMID 29432163.
- ^ Heyes, P. J.; Anastasakis, K.; de Jong, W.; van Hoesel, A.; Roebroeks, W.; Soressi, M. (2016). "Selection and Use of Manganese Dioxide by Neanderthals". Scientific Reports. 6 (1): 22,159. Bibcode:2016NatSR...622159H. doi:10.1038/srep22159. PMC 4770591. PMID 26922901.
- ^ Jaubert, J.; Verheyden, S.; Genty, D.; et al. (2016). "Early Neanderthal constructions deep in Bruniquel Cave in southwestern France". Nature. 534 (7605): 111–114. Bibcode:2016Natur.534..111J. doi:10.1038/nature18291. PMID 27251286. S2CID 205249458.
- ^ Degano, I.; Soriano, S.; Villa, P.; Pollarolo, L.; Lukejko, J. J.; Jacobs, Z.; Douka, K.; Vitagliano, S.; Tozzi, C. (2019). "Hafting of Middle Paleolithic tools in Latium (central Italy): new data from Fossellone and Sant'Agostino caves". PLOS ONE. 14 (6): e0213473. Bibcode:2019PLoSO..1413473D. doi:10.1371/journal.pone.0213473. PMC 6586293. PMID 31220106.
- ^ Sørensen, B. (2009). "Energy use by Eem Neanderthals". Journal of Archaeological Science. 36 (10): 2201–2205. doi:10.1016/j.jas.2009.06.003.
- ^ a b Collard, M.; Tarle, L.; Sandgathe, D.; Allan, A. (2016). "Faunal evidence for a difference in clothing use between Neanderthals and early modern humans in Europe". Journal of Anthropological Archaeology. 44: 235–246. doi:10.1016/j.jaa.2016.07.010. hdl:2164/9989.
- ^ a b Wales, N. (2012). "Modeling Neanderthal clothing using ethnographic analogues". Journal of Human Evolution. 63 (6): 781–795. doi:10.1016/j.jhevol.2012.08.006. PMID 23084621.
- ^ a b Carter, T.; Contreras, D. A.; Holcomb, J.; Mihailović, D. D. (2019). "Earliest occupation of the Central Aegean (Naxos), Greece: implications for hominin and Homo sapiens' behavior and dispersals". Science Advances. 5 (10): eaax0997. Bibcode:2019SciA....5..997C. doi:10.1126/sciadv.aax0997. PMC 6795523. PMID 31663021.
- ^ Johansson, S. (2015). "Language abilities in Neanderthals". Annual Review of Linguistics. 1: 311–322. doi:10.1146/annurev-linguist-030514-124945.
- ^ a b Whiting, K.; Konstantakos, L.; Sadler, G.; Gill, C. (2018). "Were Neanderthals rational? A stoic approach". Humanities. 7 (2): 39. doi:10.3390/h7020039.
- ^ Krause, J.; Lalueza-Fox, C.; Orlando, L. (2007). "The derived FOXP2 variant of modern humans was shared with Neandertals". Current Biology. 17 (21): 1, 908–1, 912. doi:10.1016/j.cub.2007.10.008. hdl:11858/00-001M-0000-000F-FED3-1. PMID 17949978. S2CID 9518208.
- ^ Mozzi, A.; Forni, D.; Clerici, M.; Pozzoli, U.; Mascheretti, S. (2016). "The evolutionary history of genes involved in spoken and written language: beyond FOXP2". Scientific Reports. 6: 22157. Bibcode:2016NatSR...622157M. doi:10.1038/srep22157. PMC 4766443. PMID 26912479.
- ^ Murphy, E.; Benítez-Burraco, A. (2017). "Paleo-oscillomics: inferring aspects of Neanderthal language abilities from gene regulation of neural oscillations" (PDF). Journal of Anthropological Sciences. 96 (96): 111–124. doi:10.4436/JASS.96010. PMID 30566085.
- ^ Lieberman, P.; Crelin, E. S. (1971). "On the speech of Neanderthal man". Linguistic Inquiry. 2 (2): 203–222. doi:10.1515/9783110819434-007. JSTOR 4177625.
- ^ Lieberman, P. (1992). "On Neanderthal speech and Neanderthal extinction". Current Anthropology. 33 (4): 409–410. doi:10.1086/204092. S2CID 144005974.
- ^ Lieberman, P. (2007). "Current views on Neanderthal speech capabilities: A reply to Boe et al. (2002)". Journal of Phonetics. 35 (4): 552–563. doi:10.1016/j.wocn.2005.07.002.
- ^ Boë, L.-J.; Heim, J.-L.; Honda, K.; Maeda, S. (2002). "The potential Neandertal vowel space was as large as that of modern humans". Journal of Phonetics. 30 (3): 465–484. doi:10.1006/jpho.2002.0170. S2CID 16685699.
- ^ Boë, L.J.; Heim, J.L.; Honda, K.; Maeda, S.; Badin, P.; Abry, C. (2007). "The vocal tract of newborn humans and Neanderthals: Acoustic capabilities and consequences for the debate on the origin of language. A reply to Lieberman". Journal of Phonetics. 35 (4): 564–581. doi:10.1016/j.wocn.2007.06.006.
- ^ Lieberman, P. (2015). "Language did not spring forth 100,000 years ago". PLOS Biology. 13 (2): e1002064. doi:10.1371/journal.pbio.1002064. PMC 4332681. PMID 25679377.
- ^ Kissel, M.; Fuentes, A. (2018). "'Behavioral modernity' as a process, not an event, in the human niche". Time and Mind. 11 (2): 163–183. doi:10.1080/1751696X.2018.1469230. S2CID 165770309.
- ^ Sterelny, K. (2011). "From hominins to humans: how sapiens became behaviourally modern". Philosophical Transactions of the Royal Society B. 366 (1566): 809–822. doi:10.1098/rstb.2010.0301. PMC 3048993. PMID 21320896.
- ^ Stringer 1993, pp. 158–60.
- ^ a b Rendu, W.; Beauval, C.; Crevecoeur, I.; Bayle, P.; Balzeau, A.; Bismuth, T.; Bourguignon, L.; Delfour, G.; Faivre, J.-P.; Lacrampe-Cuyaubère, F.; Muth, X.; Pasty, S.; Semal, P.; Tavormina, C.; Todisco, D.; Turq, A.; Maureille, B. (2016). "Let the dead speak…comments on Dibble et al.'s reply to 'Evidence supporting an intentional burial at La Chapelle-aux-Saints'". Journal of Archaeological Science. 69: 12–20. doi:10.1016/j.jas.2016.02.006.
- ^ Gargett, R. H. (1989). "Grave shortcomings: the evidence for Neandertal burial". Current Anthropology. 30 (2): 157–190. doi:10.1086/203725. S2CID 146781901.
- ^ Gargett, R. H. (1999). "Middle Palaeolithic burial is not a dead issue: the view from Qafzeh, Saint-Césaire, Kebara, Amud, and Dederiyeh". Journal of Human Evolution. 37 (1): 27–90. doi:10.1006/jhev.1999.0301. PMID 10375476. S2CID 40037076.
- ^ a b c Wunn, I. (2001). "Cave bear worship in the Paleolithic" (PDF). Cadernos do Laboratorio Xeolóxico de Laxe. 26: 457–463.
- ^ Vyshedskiy, A. (2017). "Language evolution to revolution: from a slowly developing finite communication system with many words to infinite modern language". Research Ideas and Outcomes. 5: e38546. bioRxiv 10.1101/166520. doi:10.3897/rio.5.e38546.
- ^ Rendu, W.; Beauval, C.; Crevecoeur, I.; et al. (2014). "Evidence supporting an intentional Neandertal burial at La Chapelle-aux-Saints". Proceedings of the National Academy of Sciences. 111 (1): 81–86. Bibcode:2014PNAS..111...81R. doi:10.1073/pnas.1316780110. PMC 3890882. PMID 24344286.
- ^ Dibble, H.; Aldeias, V.; Goldberg, P.; Sandgathe, D.; Steele, T. E. (2015). "A critical look at evidence from La Chapelle-aux-Saints supporting an intentional burial". Journal of Archaeological Science. 53: 649–657. doi:10.1016/j.jas.2014.04.019.
- ^ Leroi-Gourhan, A. (1975). "The flowers found with Shanidar IV, a Neanderthal burial in Iraq". Science. 190 (4214): 562–564. Bibcode:1975Sci...190..562L. doi:10.1126/science.190.4214.562. S2CID 140686473.
- ^ Solecki, R. S. (1975). "Shanidar IV: a Neanderthal flower burial in northern Iraq". Science. 190 (4217): 880–881. Bibcode:1975Sci...190..880S. doi:10.1126/science.190.4217.880. S2CID 71625677.
- ^ Sommer, J. D. (1999). "The Shanidar IV 'flower burial': a re-evaluation of Neanderthal burial ritual". Cambridge Archaeological Journal. 9 (1): 127–129. doi:10.1017/s0959774300015249.
- ^ Carroll, M. P. (1986). "The bear cult that wasn't: a study in the psychohistory of anthropology". Journal of Psychoanalytic Anthropology. 19 (1): 19–34.
- ^ Blanc, A. C. (1962). "Some evidence for the early ideologies of man". In Washburn, S. L. (ed.). Social life of early man. Routledge. pp. 124–126. ISBN 978-1-136-54361-6.
- ^ White, T. D.; Toth, N.; Chase, P. G. (1991). "The question of ritual cannibalism at Grotta Guattari". Current Anthropology. 32 (7): 118–138. doi:10.1086/203931. JSTOR 2743640. S2CID 144628134.
- ^ a b Fu, Q.; Hajdinjak, M.; Moldovan, O. T.; et al. (2015). "An early modern human from Romania with a recent Neanderthal ancestor". Nature. 524 (7564): 216–219. Bibcode:2015Natur.524..216F. doi:10.1038/nature14558. PMC 4537386. PMID 26098372.
- ^ Sankararaman, S.; Patterson, N.; Li, H.; Pääbo, S.; Reich, D; Akey, J. M. (2012). "The date of interbreeding between Neandertals and modern humans". PLOS Genetics. 8 (10): e1002947. arXiv:1208.2238. Bibcode:2012arXiv1208.2238S. doi:10.1371/journal.pgen.1002947. PMC 3464203. PMID 23055938.
- ^ a b Yang, M. A.; Malaspinas, A. S.; Durand, E. Y.; Slatkin, M. (2012). "Ancient structure in Africa unlikely to explain Neanderthal and non-African genetic similarity". Molecular Biology and Evolution. 29 (10): 2, 987–2, 995. doi:10.1093/molbev/mss117. PMC 3457770. PMID 22513287.
- ^ Sánchez-Quinto, F.; Botigué, L. R.; Civit, S.; Arenas, C.; Ávila-Arcos, M. C.; Bustamante, C. D.; Comas, D.; Lalueza-Fox, C.; Caramelli, D. (2012). "North African populations carry the signature of admixture with Neandertals". PLOS ONE. 7 (10): e47765. Bibcode:2012PLoSO...747765S. doi:10.1371/journal.pone.0047765. PMC 3474783. PMID 23082212.
- ^ Yotova, V.; Lefebvre, J.-F.; Moreau, C.; et al. (2011). "An X-linked haplotype of Neandertal origin is present among all non-African populations". Molecular Biology and Evolution. 28 (7): 1957–1962. doi:10.1093/molbev/msr024. PMID 21266489.
- ^ Fu, Q.; Li, H.; Moorjani, P.; et al. (2014). "Genome sequence of a 45,000-year-old modern human from western Siberia". Nature. 514 (7523): 445–449. Bibcode:2014Natur.514..445F. doi:10.1038/nature13810. PMC 4753769. PMID 25341783.
- ^ Lohse, K.; Frantz, L. A. F. (2013). "Maximum likelihood evidence for Neandertal admixture in Eurasian populations from three genomes". Populations and Evolution. 1307: 8263. arXiv:1307.8263. Bibcode:2013arXiv1307.8263L.
- ^ Prüfer, K.; de Filippo, C.; Grote, S.; Mafessoni, F.; Korlević, P.; Hajdinjak, M.; et al. (2017). "A high-coverage Neandertal genome from Vindija Cave in Croatia". Science. 358 (6363): 655–658. Bibcode:2017Sci...358..655P. doi:10.1126/science.aao1887. PMC 6185897. PMID 28982794.
- ^ Chen, L.; Wolf, A. B.; Fu, W.; Akey, J. M. (2020). "Identifying and Interpreting Apparent Neanderthal Ancestry in African Individuals". Cell. 180 (4): 677–687.e16. doi:10.1016/j.cell.2020.01.012. PMID 32004458. S2CID 210955842.
- ^ Pääbo, S. (2015). "The diverse origins of the human gene pool". Nature Reviews Genetics. 16 (6): 313–314. doi:10.1038/nrg3954. PMID 25982166. S2CID 5628263.
- ^ Enard, D.; Petrov, D. A. (2018). "Evidence that RNA viruses drove of adaptive introgression between Neanderthals and modern humans". Cell. 175 (2): 360–371. doi:10.1016/j.cell.2018.08.034. PMC 6176737. PMID 30290142.
- ^ Taskent, R. O.; Alioglu, N. D.; Fer, E.; et al. (2017). "Variation and functional impact of Neanderthal ancestry in Western Asia". Genome Biology and Evolution. 9 (12): 3516–3624. doi:10.1093/gbe/evx216. PMC 5751057. PMID 29040546.
- ^ Zorina-Lichtenwalter, K.; Lichtenwalter, R. N.; Zaykin, D. V.; et al. (2019). "A study in scarlet: MC1R as the main predictor of red hair and exemplar of the flip-flop effect". Human Molecular Genetics. 28 (12): 2093–2106. doi:10.1093/hmg/ddz018. PMC 6548228. PMID 30657907.
- ^ Ding, Q.; Hu, Y.; Xu, S.; Wang, C.-C.; Li, H.; Zhang, R.; Yan, S.; Wang, J.; Jin, L. (2014). "Neanderthal origin of the haplotypes carrying the functional variant Val92Met in the MC1R in modern humans". Molecular Biology and Evolution. 31 (8): 1994–2003. doi:10.1093/molbev/msu180. PMID 24916031.
- ^ a b Mendez, F. L.; Watkins, J. C.; Hammer, M. F. (2013). "Neandertal origin of genetic variation at the cluster of OAS immunity genes" (PDF). Molecular Biology and Evolution. 30 (4): 798–801. doi:10.1093/molbev/mst004. PMID 23315957. S2CID 2839679. Archived from the original (PDF) on February 23, 2019.
- ^ a b Mendez, F. L.; Watkins, J. C.; Hammer, M. F. (2012). "A haplotype at STAT2 introgressed from Neanderthals and serves as a candidate of positive selection in Papua New Guinea". American Journal of Human Genetics. 91 (2): 265–274. doi:10.1016/j.ajhg.2012.06.015. PMC 3415544. PMID 22883142.
- ^ a b Dannemann, M.; Andrés, A. A.; Kelso, J. (2016). "Introgression of Neandertal- and Denisovan-like haplotypes contributes to adaptive variation in human toll-like receptors". American Journal of Human Genetics. 98 (1): 22–33. doi:10.1016/j.ajhg.2015.11.015. PMC 4716682. PMID 26748514.
- ^ Gregory, M. D.; Kippenhan, J. S.; Eisenberg, D. P.; et al. (2017). "Neanderthal-derived genetic variation shapes modern human cranium and brain". Scientific Reports. 7 (1): 6308. Bibcode:2017NatSR...7.6308G. doi:10.1038/s41598-017-06587-0. PMC 5524936. PMID 28740249.
- ^ a b Luo, S.; Valencia, C. A.; Zhang, J.; Lee, N.-C.; Slone, J.; Gui, B.; Wang, X.; Li, Z.; Dell, S.; Brown, J.; Chen, S. M.; Chien, Y.-H.; Hwu, W.-L.; Fan, P.-C.; Wong, L.-J.; Atwal, P. S.; Huang, T. (2018). "Biparental inheritance of mitochondrial DNA in humans". Proceedings of the National Academy of Sciences. 115 (51): 13039–13044. doi:10.1073/pnas.1810946115. PMC 6304937. PMID 30478036.
- ^ Mason, P. H.; Short, R. V. (2011). "Neanderthal-human hybrids". Hypothesis. 9: e1.
- ^ Mendez, F. L.; Poznik, G. D.; Castellano, S.; Bustamante, C. D. (2016). "The divergence of Neandertal and modern human Y chromosomes". American Journal of Human Genetics. 98 (4): 728–734. doi:10.1016/j.ajhg.2016.02.023. PMC 4833433. PMID 27058445.
- ^ Sankararaman, S.; Patterson, N.; Li, H.; Pääbo, S.; Reich, D. (2012). "The date of interbreeding between Neandertals and modern humans". PLOS Genetics. 8 (10): e1002947. arXiv:1208.2238. Bibcode:2012arXiv1208.2238S. doi:10.1371/journal.pgen.1002947. PMC 3464203. PMID 23055938.
- ^ Hershkovitz, I.; Weber, G. W.; Quam, R.; Duval, M.; Grün, R. (2018). "The earliest modern humans outside Africa". Science. 359 (6374): 459. Bibcode:2018Sci...359..456H. doi:10.1126/science.aap8369. hdl:10072/372670. PMID 29371468.
- ^ McDermott, F.; Grün, R.; Stringer, C. B.; Hawkesworth, C. J. (1993). "Mass-spectrometric U-series dates for Israeli Neanderthal/early modern hominid sites". Nature. 363 (6426): 252–255. Bibcode:1993Natur.363..252M. doi:10.1038/363252a0. PMID 8387643. S2CID 4234812.
- ^ Pagani, L. (2016). "Genomic analyses inform on migration events during the peopling of Eurasia". Nature. 538 (7624): 238–242. Bibcode:2016Natur.538..238P. doi:10.1038/nature19792. PMC 5164938. PMID 27654910.
- ^ Lowery, R. K.; Uribe, G.; Jimenez, E. B.; Weiss, M. A.; Herrera, K. J.; Regueiro, M.; Herrera, R. J. (2013). "Neanderthal and Denisova genetic affinities with contemporary humans: Introgression versus common ancestral polymorphisms". Gene. 530 (1): 83–94. doi:10.1016/j.gene.2013.06.005. PMID 23872234.
- ^ Stringer, C. (2012). "Evolution: what makes a modern human". Nature. 485 (7396): 33–35. Bibcode:2012Natur.485...33S. doi:10.1038/485033a. PMID 22552077. S2CID 4420496.
- ^ Pääbo, S. (2014). "A mitochondrial genome sequence of a hominin from Sima de los Huesos" (PDF). Nature. 505 (7483): 403–406. Bibcode:2014Natur.505..403M. doi:10.1038/nature12788. PMID 24305051. S2CID 4456221.
- ^ Warren, Matthew (2018). "Mum's a Neanderthal, dad's a Denisovan: First discovery of an ancient-human hybrid". Nature News. 560 (7719): 417–418. Bibcode:2018Natur.560..417W. doi:10.1038/d41586-018-06004-0. PMID 30135540.
- ^ Agusti, J.; Rubio-Campillo, X. (2017). "Were Neanderthals responsible for their own extinction?". Quaternary International. 431: 232–237. Bibcode:2017QuInt.431..232A. doi:10.1016/j.quaint.2016.02.017.
- ^ a b Zilhão, J.; Anesin, D.; Aubry, T.; et al. (2017). "Precise dating of the Middle-to-Upper Paleolithic transition in Murcia (Spain) supports late Neandertal persistence in Iberia". Heliyon. 3 (1): E00435. doi:10.1016/j.heliyon.2017.e00435. PMC 5696381. PMID 29188235.
- ^ Morales, J. I; Cebrià, A.; Burguet-Coca, A.; et al. (2019). "The Middle-to-Upper Paleolithic transition occupations from Cova Foradada (Calafell, NE Iberia)". PLOS ONE. 14 (5): e0215832. Bibcode:2019PLoSO..1415832M. doi:10.1371/journal.pone.0215832. PMC 6522054. PMID 31095578.
- ^ Cortés-Sánches, M.; Jiménez-Espejo, F. J.; Simón-Vallejo, M. D.; Stringer, C.; et al. (2019). "An early Aurignacian arrival in southwestern Europe". Nature Ecology & Evolution. 3 (2): 207–212. doi:10.1038/s41559-018-0753-6. PMID 30664696. S2CID 58571988.
- ^ Anderson, L.; Reynolds, N.; Teyssandier, N. (2019). "No reliable evidence for a very early Aurignacian in Southern Iberia". Nature Ecology and Evolution. 3 (5): 713. doi:10.1038/s41559-019-0885-3. PMID 30988496. S2CID 115153460.
- ^ de la Peña, P. (2019). "Dating on its own cannot resolve hominin occupation patterns". Nature Ecology and Evolution. 3 (5): 712. doi:10.1038/s41559-019-0886-2. PMID 30988497. S2CID 116861216.
- ^ Cortés-Sánches, M.; Jiménez-Espejo, F. J.; Simón-Vallejo, M. D.; Stringer, C.; et al. (2019). "Reply to 'Dating on its own cannot resolve hominin occupation patterns' and 'No reliable evidence for a very early Aurignacian in Southern Iberia'". Nature Ecology and Evolution. 3 (5): 714–715. doi:10.1038/s41559-019-0887-1. PMID 30988498. S2CID 115153407.
- ^ Fontugne, M.; Reyss, J. L.; Ruis, C. B.; Lara, P. M. (1995). "The Mousterian site of Zafarraya (Granada, Spain): dating and implications on the palaeolithic peopling processes of Western Europe". Comptes Rendus de l'Académie des Sciences. 321 (10): 931–937.
- ^ Finlayson, C.; Pacheco, F. G. (2006). "Late survival of Neanderthals at the southernmost extreme of Europe". Nature. 443 (7, 113): 850–853. Bibcode:2006Natur.443..850F. doi:10.1038/nature05195. PMID 16971951. S2CID 4411186.
- ^ Pavlov, P.; Svendsen, J. I.; Indrelid, S. (2001). "Human presence in the European Arctic nearly 40,000 years ago". Nature. 413 (6, 851): 64–67. Bibcode:2001Natur.413...64P. doi:10.1038/35092552. PMID 11544525. S2CID 1986562.
- ^ Hublin, J.-J.; Sirakov, N.; et al. (2020). "Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria" (PDF). Nature. 581 (7808): 299–302. Bibcode:2020Natur.581..299H. doi:10.1038/s41586-020-2259-z. PMID 32433609. S2CID 218592678.
- ^ Benazzi, S.; Douka, K.; Fornai, C.; Bauer, C.C.; Kullmer, O.; Svoboda, J.Í.; Pap, I.; Mallegni, F.; Bayle, P.; Coquerelle, M.; Condemi, S.; Ronchitelli, A.; Harvati, K.; Weber, G.W. (2011). "Early dispersal of modern humans in Europe and implications for Neanderthal behaviour". Nature. 479 (7374): 525–528. Bibcode:2011Natur.479..525B. doi:10.1038/nature10617. PMID 22048311. S2CID 205226924.
- ^ Higham, T.; Compton, T.; Stringer, C.; Jacobi, R.; Shapiro, B.; Trinkaus, E.; Chandler, B.; Gröning, F.; Collins, C.; Hillson, S.; o’Higgins, P.; Fitzgerald, C.; Fagan, M. (2011). "The earliest evidence for anatomically modern humans in northwestern Europe". Nature. 479 (7374): 521–524. Bibcode:2011Natur.479..521H. doi:10.1038/nature10484. PMID 22048314. S2CID 4374023.
- ^ Harvati, K.; et al. (2019). "Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia". Nature. 571 (7766): 500–504. doi:10.1038/s41586-019-1376-z. PMID 31292546. S2CID 195873640.
- ^ de Lumley, M.-A.; Guipert, G.; de Lumley, H.; Protopapa, N.; Pitsios, T. (2020). "Apidima 1 and Apidima 2: two anteneandertal skulls in the Peloponnese, Greece". L'Anthropologie. 124 (1): 102743. doi:10.1016/j.anthro.2019.102743. S2CID 216449347.
- ^ Staubwasser, M.; Drăgușin, V.; Onac, B. P. (2018). "Impact of climate change on the transition of Neanderthals to modern humans in Europe". Proceedings of the National Academy of Sciences. 115 (37): 9116–9121. Bibcode:2018PNAS..115.9116S. doi:10.1073/pnas.1808647115. PMC 6140518. PMID 30150388.
- ^ Degioanni, A.; Bonenfant, C.; Cabut, S.; Condemi, S. (2019). "Living on the edge: Was demographic weakness the cause of Neanderthal demise?". PLOS ONE. 14 (5): e0216742. Bibcode:2019PLoSO..1416742D. doi:10.1371/journal.pone.0216742. PMC 6541251. PMID 31141515.
- ^ Marti, A.; Folch, A.; Costa, A.; Engwell, S. (2016). "Reconstructing the plinian and co-ignimbrite sources of large volcanic eruptions: A novel approach for the Campanian Ignimbrite". Scientific Reports. 6 (21220): 1–11. Bibcode:2016NatSR...621220M. doi:10.1038/srep21220. PMC 4756320. PMID 26883449.
- ^ Liberski, P. (2013). "Kuru: a journey back in time from Papua New Guinea to the Neanderthals' extinction". Pathogens. 2 (3): 472–505. doi:10.3390/pathogens2030472. PMC 4235695. PMID 25437203.
General bibliography[]
- Brown, K.; Fa, D. A.; Finlayson, G.; Finlayson, C. (2011). "Small game and marine resource exploitation by Neanderthals: the evidence from Gibraltar". Trekking the shore: changing coastlines and the antiquity of coastal settlement. Interdisciplinary contributions to archaeology. Springer. ISBN 978-1-4419-8218-6.
- Finlayson, C. (2019). The smart Neanderthal: bird catching, cave art, and the cognitive revolution. Oxford University Press. ISBN 978-0-19-251812-5.
- Papagianni, D.; Morse, M. A. (2013). "Still with us?". Neanderthals rediscovered: how modern science is rewriting their story. Thames and Hudson. ISBN 978-0-500-77311-6.
- Reich, D. (2018). "Encounters with Neanderthals". Who we are and how we got here: ancient DNA and the new science of the human past. Oxford University Press. ISBN 978-0-19-882125-0.
- Shipman, P. (2015). "How humans and their dogs drove Neanderthals to extinction". The invaders: how humans and their dogs drove Neanderthals to extinction. Harvard University Press. ISBN 978-0-674-42538-5. JSTOR j.ctvjf9zbs.
- Stringer, C.; Gamble, C. (1993). In search of the Neanderthals. Thames and Hudson. ISBN 978-0-500-05070-5.
- Sykes, Rebecca Wragg (2020). Kindred: Neanderthal life, love, death and art. London, UK: Bloomsbury Sigma. ISBN 978-1-4729-3749-0.
- Tattersall, I. (2015). "Neanderthals, DNA, and creativity". The strange case of the Rickety Cossack: and other cautionary tales from human evolution. St. Martin's Publishing Group. ISBN 978-1-4668-7943-0.
External links[]
| Wikimedia Commons has media related to Homo neanderthalensis. |
| Wikibooks has a book on the topic of: Introduction to Paleoanthropology |
| Wikispecies has information related to Homo neanderthalensis. |
| Wikisource has the text of the 1905 New International Encyclopedia article "Neanderthal Man". |
| Wikinews has related news: |
- "Homo neanderthalensis". The Smithsonian Institution. February 14, 2010.
- Human Timeline (Interactive) – Smithsonian, National Museum of Natural History (August 2016).
- "Neanderthal DNA". International Society of Genetic Genealogy. Archived from the original on June 17, 2006.: Includes Neanderthal mtDNA sequences
- GenBank records for H. s. neanderthalensis maintained by the National Center for Biotechnology Information (NCBI)
Human evolution | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines |
| ||||||||||||||||||||||||||
| Others |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
Authority control | |
|---|---|
| General | |
| National libraries | |
| Other | |
- Neanderthals
- Fossil taxa described in 1864
- Stone Age Europe
- Stone Age Asia