Redfield ratio
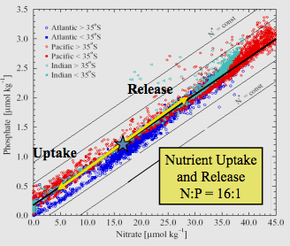
Redfield ratio or Redfield stoichiometry is the consistent atomic ratio of carbon, nitrogen and phosphorus found in marine phytoplankton and throughout the deep oceans.
The term is named for American oceanographer Alfred C. Redfield who in 1934 first described the relatively consistent ratio of nutrients in marine biomass samples collected across several voyages on board the research vessel Atlantis, and empirically found the ratio to be C:N:P = 106:16:1.[1] While deviations from the canonical 106:16:1 ratio have been found depending on phytoplankton species and the study area, the Redfield ratio has remained an important reference to oceanographers studying nutrient limitation. A 2014 paper summarizing a large data set of nutrient measurements across all major ocean regions spanning from 1970 to 2010 reported the global median C:N:P to be 163:22:1.[2]
Discovery[]
| Part of a series on the |
| Carbon cycle |
|---|
 |
For his 1934 paper, Alfred Redfield analyzed nitrate and phosphate data for the Atlantic, Indian, Pacific oceans and Barents Sea.[1] As a Harvard physiologist, Redfield participated in several voyages on board the research vessel Atlantis, analyzing data for C, N, and P content in marine plankton, and referenced data collected by other researchers as early as 1898.
Redfield’s analysis of the empirical data led to him to discover that across and within the three oceans and Barents Sea, seawater had an N:P atomic ratio near 20:1 (later corrected to 16:1), and was very similar to the average N:P of phytoplankton.
To explain this phenomena, Redfield initially proposed two mutually non-exclusive mechanisms:
I) The N:P in plankton tends towards the N:P composition of seawater. Specifically, phytoplankton species with different N and P requirements compete within the same medium and come to reflect the nutrient composition of the seawater.[1]
II) An equilibrium between seawater and planktonic nutrient pools is maintained through biotic feedback mechanisms.[1][3] Redfield proposed a thermostat like scenario in which the activities of nitrogen fixers and denitrifiers keep the nitrate to phosphate ratio in the seawater near the requirements in the protoplasm.[4] Considering that at the time little was known about the composition of “protoplasm", or the bulk composition of phytoplankton, Redfield did not attempt to explain why its N:P ratio should be approximately 16:1.
In 1958, almost a quarter century after first discovering the ratios, Redfield leaned toward the latter mechanism in his manuscript, The Biological Control of Chemical Factors in the Environment.[3] Redfield proposed that the ratio of nitrogen to phosphorus in plankton resulted in the global ocean having a remarkably similar ratio of dissolved nitrate to phosphate (16:1). He considered how the cycles of not just N and P but also C and O could interact to result in this match.
Explanation[]
Redfield discovered the remarkable congruence between the chemistry of the deep ocean and the chemistry of living things such as phytoplankton in the surface ocean. Both have N:P ratios of about 16:1 in terms of atoms. When nutrients are not limiting, the molar elemental ratio C:N:P in most phytoplankton is 106:16:1. Redfield thought it wasn't purely coincidental that the vast oceans would have a chemistry perfectly suited to the requirements of living organisms.
Laboratory experiments under controlled chemical conditions have found that phytoplankton biomass will conform to the Redfield Ratio even when environmental nutrient levels exceed them, suggesting that ecological adaptation to oceanic nutrient ratios is not the only governing mechanism (contrary to one of the mechanisms initially proposed by Redfield).[5] However, subsequent modeling of feedback mechanisms, specifically nitrate-phosphorus coupling fluxes, do support his proposed mechanism of biotic feedback equilibrium, though these results are confounded by limitations in our current understanding of nutrient fluxes.[6]
In the ocean a large portion of the biomass is found to be nitrogen-rich plankton. Many of these plankton are consumed by other plankton biomass which have similar chemical compositions. This results in a similar nitrogen to phosphorus ratio, on average, for all the plankton throughout the world’s ocean, empirically found to average approximately 16:1. When these organisms sink into the ocean interior, their biomass is consumed by bacteria that, in aerobic conditions, oxidize the organic matter to form dissolved inorganic nutrients, mainly carbon dioxide, nitrate, and phosphate.
That the nitrate to phosphate ratio in the interior of all of the major ocean basins is highly similar is possibly due to the residence times of these elements in the ocean relative to the oceans circulation time, roughly 100 000 years for phosphorus and 2000 years for nitrogen.[7] The fact that the residence times of these elements are greater than the mixing times of the oceans (~ 1000 years)[8] can result in the ratio of nitrate to phosphate in the ocean interior remaining fairly uniform.
While such arguments can potentially explain why the ratios are fairly constant, they do not address the question why the N:P ratio is nearly 16 and not some other number.
Uses[]
The research that resulted in this ratio has become a fundamental feature in the understanding of the biogeochemical cycles of the oceans, and one of the key tenets of biogeochemistry. The Redfield ratio is instrumental in estimating carbon and nutrient fluxes in global circulation models. They also help in determining which nutrients are limiting in a localized system, if there is a limiting nutrient. The ratio can also be used to understand the formation of phytoplankton blooms and subsequently hypoxia by comparing the ratio between different regions, such as a comparison of the Redfield Ratio of the Mississippi River to the ratio of the northern Gulf of Mexico.[9] Controlling N:P could be a means for sustainable reservoir management.[10]
Deviations from the canonical Redfield ratio[]
The Redfield ratio was initially derived empirically from measurements of the elemental composition of plankton in addition to the nitrate and phosphate content of seawater collected from a few stations in the Atlantic Ocean. This was later supported by hundreds of independent measurements. However, looking at the composition of individual species of phytoplankton grown under nitrogen or phosphorus limitation shows that this nitrogen to phosphorus ratio can vary anywhere from 6:1 to 60:1. While understanding this problem, Redfield never attempted to explain it with the exception of noting that the N:P ratio of inorganic nutrients in the ocean interior was an average with small scale variability to be expected.
Although the Redfield ratio is remarkably stable in the deep ocean, phytoplankton may have large variations in the C:N:P composition, and their life strategy play a role in the C:N:P ratio, which has made some researchers speculate that the Redfield ratio perhaps is a general average rather than specific requirement for phytoplankton growth.[11] However, the Redfield ratio was recently found to be related to a homeostatic protein-to-rRNA ratio fundamentally present in both prokaryotes and eukaryotes.[12] Furthermore, the Redfield ratio has been shown to vary at different spatial scales as well as average slightly higher (166:20:1) than Redfield's original estimate.[13] In some ecosystems, the Redfield Ratio has also been shown to vary significantly by the dominant phytoplankton taxa present in an ecosystem, even in systems with abundant nutrients. Consequently, the system-specific Redfield Ratio could serve as a proxy for plankton community structure.[14]
Despite reports that the elemental composition of organisms such as marine phytoplankton in an oceanic region do not conform to the canonical Redfield ratio, the fundamental concept of this ratio remains valid and useful. In 2014, an article was released in the Scientific Data journal, which aggregated Redfield ratios measurements from observational cruises around the world from 1970 to 2010. This article provides a large database that can be used to study the evolution of particular phosphorus, carbon and nitrogen across sea stations and time.[2]
Extended Redfield ratio[]
Some feel that there are other elements, such as potassium, sulfur, zinc, copper, and iron which are also important in the ocean chemistry.[15]
In particular, iron (Fe) was considered of great importance as early biological oceanographers hypothesized that iron may also be a limiting factor for primary production in the ocean.[16] As a result an extended Redfield ratio was developed to include this as part of this balance. This new stoichiometric ratio states that the ratio should be 106 C:16 N:1 P:0.1-0.001 Fe. The large variation for Fe is a result of the significant obstacle of ships and scientific equipment contaminating any samples collected at sea with excess Fe.[17] It was this contamination that resulted in early evidence suggesting that iron concentrations were high and not a limiting factor in marine primary production.
Diatoms need, among other nutrients, silicic acid to create biogenic silica for their frustules (cell walls). As a result of this, the Redfield-Brzezinski nutrient ratio was proposed for diatoms and stated to be C:Si:N:P = 106:15:16:1.[18] Extending beyond primary production itself, the oxygen consumed by aerobic respiration of phytoplankton biomass has also been shown to follow a predictable proportion to other elements. The O2:C ratio has been measured at 138:106.[6]
See also[]
References[]
- ^ Jump up to: a b c d Redfield, AC (1934). "On the proportions of organic derivatives in sea water and their relation to the composition of plankton" (PDF). James Johnstone Memorial Volume: 176. Retrieved March 1, 2019.
- ^ Jump up to: a b Martiny, Adam C.; Vrugt, Jasper A.; Lomas, Michael W. (9 December 2014). "Concentrations and ratios of particulate organic carbon, nitrogen, and phosphorus in the global ocean". Scientific Data. 1 (1): 140048. doi:10.1038/sdata.2014.48. PMC 4421931. PMID 25977799.
- ^ Jump up to: a b REDFIELD, ALFRED C. (1958). "The Biological Control of Chemical Factors in the Environment". American Scientist. 46 (3): 205–221, 230A. ISSN 0003-0996. JSTOR 27827150. PMID 24545739.
- ^ Gruber, Nicolas; Sarmiento, Jorge L. (1997). "Global patterns of marine nitrogen fixation and denitrification". Global Biogeochemical Cycles. 11 (2): 235–266. Bibcode:1997GBioC..11..235G. doi:10.1029/97GB00077. ISSN 1944-9224.
- ^ Goldman, Joel C. (1986). "On phytoplankton growth rates and particulate C: N: P ratios at low light1". Limnology and Oceanography. 31 (6): 1358–1363. Bibcode:1986LimOc..31.1358G. doi:10.4319/lo.1986.31.6.1358. ISSN 1939-5590.
- ^ Jump up to: a b Lenton, Timothy M.; Watson, Andrew J. (2000). "Redfield revisited: 1. Regulation of nitrate, phosphate, and oxygen in the ocean". Global Biogeochemical Cycles. 14 (1): 225–248. Bibcode:2000GBioC..14..225L. doi:10.1029/1999GB900065. ISSN 1944-9224.
- ^ http://lgmacweb.env.uea.ac.uk/green_ocean/positions/diazotroph.shtml
- ^ "Chemical Sensor Group". 2015-08-20.
- ^ Dodds, Walter (2006). "Nutrients and the "dead zone" : the link between nutrient ratios and dissolved oxygens in the Northern Gulf of Mexico". Frontiers in Ecology and the Environment. 4 (4): 211–217. doi:10.1890/1540-9295(2006)004[0211:NATDZT]2.0.CO;2.
- ^ Levich, A. P. (1 February 1996). "The role of nitrogen-phosphorus ratio in selecting for dominance of phytoplankton by cyanobacteria or green algae and its application to reservoir management". Journal of Aquatic Ecosystem Health. 5 (1): 55–61. doi:10.1007/BF00691729. ISSN 1573-5141. S2CID 84469052.
- ^ Arrigo, KR (2005). "Marine microorganisms and global nutrient cycles". Nature. 437 (7057): 349–55. Bibcode:2005Natur.437..349A. doi:10.1038/nature04159. PMID 16163345. S2CID 62781480.
- ^ Loladze, Irakli; Elser, James J. (2011). "The origins of the Redfield nitrogen-to-phosphorus ratio are in a homoeostatic protein-to-rRNA ratio". Ecology Letters. 14 (3): 244–250. doi:10.1111/j.1461-0248.2010.01577.x. ISSN 1461-0248. PMID 21244593.
- ^ Sterner, Robert W.; Andersen, Tom; Elser, James J.; Hessen, Dag O.; Hood, James M.; McCauley, Edward; Urabe, Jotaro (2008). "Scale-dependent carbon:nitrogen:phosphorus seston stoichiometry in marine and freshwaters". Limnology and Oceanography. 53 (3): 1169–1180. Bibcode:2008LimOc..53.1169S. doi:10.4319/lo.2008.53.3.1169. ISSN 1939-5590.
- ^ Arrigo, Kevin R.; Dunbar, Robert B.; Lizotte, Michael P.; Robinson, D. H. (2002). "Taxon-specific differences in C/P and N/P drawdown for phytoplankton in the Ross Sea, Antarctica". Geophysical Research Letters. 29 (19): 44–1–44-4. Bibcode:2002GeoRL..29.1938A. doi:10.1029/2002GL015277. ISSN 1944-8007.
- ^ Benner, R; Pakulski, JD; McCarthy, M; Hedges, JI; Hatcher, PG (1992). "Bulk chemical characteristics of dissolved organic matter in the ocean". Science. 255 (5051): 1561–4. Bibcode:1992Sci...255.1561B. doi:10.1126/science.255.5051.1561. PMID 17820170. S2CID 28604425.
- ^ Tortell, PD; Maldonado, MT; Granger, J; Price, NM (1999). "Marine bacteria and biogeochemical cycling of iron in the oceans". Microbiology Ecology. 29 (1): 1. doi:10.1111/j.1574-6941.1999.tb00593.x.
- ^ Broecker, WS; Peng, T; Beng, Z (1982). Tracers in the Sea. Lamont-Doherty Geological Observatory.
- ^ Brzezinski, Mark A. (1985). "THE Si:C:N RATIO OF MARINE DIATOMS: INTERSPECIFIC VARIABILITY AND THE EFFECT OF SOME ENVIRONMENTAL VARIABLES1". Journal of Phycology. 21 (3): 347–357. doi:10.1111/j.0022-3646.1985.00347.x. ISSN 1529-8817. S2CID 86568341.
- Johnson, Zackary. "Biogeochemistry IV." University of Hawaii School of Ocean and Earth Science and Technology. Web. <http://www.soest.hawaii.edu/oceanography/zij/ocn621/OCN621-20060215-biogeochemistry.pdf>.
- Lentz, Jennifer. "Nutrient Stoichiometry - Redfield Ratios." LSU School of the Coast and Environment, 2010. Web. <http://www.sce.lsu.edu/cego/Documents/Reviews/Oceanography/Nutrient_Stoichiometry.pdf>.
- P.G. Falkowski, and C.S. Davis. "MARINE BIOGEOCHEMISTRY: ON REDFIELD RATIOS." ScienceWeek. Nature, 2004. Web. <http://scienceweek.com/2004/sa041119-5.htm>.
| show Authority control |
|---|
- Ecology
- Ratios